Характеризація звуків courtship видів підгрупи fasciola (Diptera, Drosophilidae, група Drosophila repleta): аналізи міжвидових та міжпопуляційних взаємодій
Машинний переклад
Оригінальна стаття написана мовою EN (посилання для прочитання) .
Анотація
Метою цієї роботи було охарактеризувати шаблон чоловічої пісні під час courtship різних видів підгрупи fasciola та визначити рівень варіації як всередині, так і між видами. Аналізованими параметрами були інтервал між імпульсами (PI), інтервал між імпульсами (IPI) та частота імпульсу (IF). Було проаналізовано шість різних видів: D. coroica (три популяції), D. ellisoni, D. fascioloides, D. moju, D. onca та D. rosinae (по одній популяції кожен). Між цими шістьма видами були виявлені значні відмінності за цими трьома параметрами пісні courtship. IPI був найбільш змінним параметром серед цих видів, що свідчить про те, що цей параметр важливий для дискримінації самок. Чотири різні гіпотези можуть пояснити цю варіацію: 1. різні селекційні тиски з відсутністю генетичного потоку; 2. інтраспецифічний статевий відбір; 3. симпатричні ефекти на еволюцію пісні; та 4. генетичний дрейф. PI був єдиним параметром, який суттєво відрізнявся між трьома популяціями D. coroica. Низька варіабельність між популяціями одного й того ж виду вже спостерігалася для інших підгруп і може бути пояснена наступними гіпотезами: сильний відбір, що діє на параметри пісні, генетичний потік або нещодавня колонізація з загального джерела. Додаткові дослідження пісні courtship інших видів підгрупи fasciola, а також для інших підгруп групи repleta, і дослідження, що використовують молекулярні маркери, які зосереджуються на генетичних основах відмінностей між цими видами в пісні courtship, дозволять нам оцінити асоціацію пісні courtship і статевої ізоляції в цих видах, а також допоможуть зрозуміти еволюцію цих поведінкових відмінностей.
Вступ
Звуки залицяння
Повадки залицяння мух роду Drosophila є важливими для розпізнавання видів. Після того, як Шорі (1962) показав, що самці D. melanogaster видають звуки під час сексуального залицяння, було проведено багато досліджень з різними групами та підгрупами роду Drosophila, що описують специфічну природу типів звуків. Ці дослідження надали докази того, що звук виконує функцію як сексуального стимулятора, так і в сексуальній ізоляції. Ці звуки значно варіюються серед різних видів груп affinis (Chang & Miller, 1978), melanogaster (Cowling & Burnet, 1981), virilis (Hoikkala et al., 1982; Hoikkala & Isoherranen, 1997), mercatorum (Ikeda & Maruo, 1982), repleta (Ewing & Miyan, 1986), auraria (Tomaru & Oguma, 1994), planitibia (Hoikkala et al., 1994), willistoni (Ritchie & Gleason, 1995), quinaria (Neems et al., 1997) та obscura (Noor & Aquadro, 1998).
Міжвидові відмінності в Drosophila звуках залицяння характеризуються одним або кількома звуковими параметрами, такими як: інтерпульсний інтервал (IPI), який є часом від початку одного імпульсу до початку наступного; інтраімпульсний інтервал (PI), який є тривалістю імпульсу; інтраімпульсна частота (IF), яка є акустично найсильнішою частотою в межах імпульсу; кількість імпульсів за серію; та тривалість кожного інтервалу в межах серії (Hoikkala & Lumme, 1987; Wheeler et al., 1988). Хоча ще не було чітко встановлено, які звукові параметри надають специфічність, є докази того, що IPI є важливим. Згідно з Ritchie & Kyriacou (1996), IPI звуків залицяння Drosophila варіює дуже мало в природних популяціях і є важливим для репродуктивного успіху самців.
Проте очевидно, що багато видів Drosophila випускають складні звуки з різними характеристиками, один або більше з яких можуть бути корисними для ідентифікації видів. Ця специфічність у звуках залицяння Drosophila може бути пояснена відбором для розрізнення між видами (Tomaru & Oguma, 1994) різних типів звуків, які могли еволюціонувати через інтраспецифічний сексуальний відбір (Ritchie & Gleason, 1995). Також різні типи звуків, що зустрічаються в різних видах Drosophila , можуть просто відображати філогенетичні розбіжності.
Підгрупа фасціоли
Види підгрупи фасціола зустрічаються у вологих лісах, переважно в Центральній та Південній Америці, а також на Карибських островах. У Бразилії їх збирали в Атлантичному тропічному лісі, а також у лісах штатів Сан-Паулу, Парана, Мату-Гросу-ду-Сул і Баїя (Вілела, 1983; Вілела та ін., 1983; Тідон-Склорз & Сене, 1992).
Більшість досліджень цієї підгрупи були цитологічними і проводилися на початку 60-х років, показуючи, що види в цій групі відрізняються від усіх інших видів групи реплета наявністю гомозиготних інверсій: 2o2, 2e3 та 2l3. Вассерман (1982) запропонував спільне походження для підгруп фасціола та муллері, оскільки вони мають спільну інверсію 3c. Однак нещодавно Дініз (1998) показав, що інверсія 3c підгрупи фасціола відрізняється від тієї, що знайдена в підгрупі муллері, і запропонував, що підгрупа фасціола походить безпосередньо від примітивної I і більше не може вважатися пов'язаною з підгрупою муллері. Кун та ін. (1995) провели каріотипічне дослідження деяких видів підгрупи фасціола і виявили міжвидову варіацію в розмірах мікрохромосом і в числі хромосом.
Згідно з Вассерманом (1992), підгрупа fasciola складається з дев'яти видів: D. pictilis та D. pictura, які формують комплекс pictilis; D. paraguttata; D. mojuoides та D. moju, які формують комплекс moju; а також D. fulvalineata, D. fasciola, D. coroica та D. fascioloides, які не входять до жодного комплексу. Вілела (1983), вивчаючи морфологію чоловічих статевих органів, визначив, що вид D. fascioloides, вивчений Вассерманом (1962), насправді не належав до цього виду, і перейменував його на D. ellisoni. Він також додав види D. linearepleta та D. onca до підгрупи fasciola і описав D. carolinae, D. hermionae, D. ivai, D. querubimae, D. rosinae та D. senei як нові види, збільшивши кількість видів у цій підгрупі до 18.
У цій роботі ми описуємо звукові патерни шлюбного поведінки шести видів підгрупи fasciola: D. ellisoni, D. fascioloides, D. moju, D. onca, D. rosinae та D. coroica. Окрім міжвидових аналізів, ми також вивчили відмінності у звукових патернах трьох різних популяцій D. coroica.
Матеріали та методи
Встановлення та підтримка ізоліній
Ізолінії були отримані від окремих самок, які вже спарувалися в природі. Ці ізолінії були ідентифіковані через морфологічний аналіз геніталій самців F1 (Vilela, 1983). Личинки третього віку підгрупи fasciola зазвичай намагаються вилізти з культури. Щоб уникнути цієї проблеми, культури були відкриті та розміщені на вологому піску в пластикових пляшках, щоб личинки могли перетворитися в лялечок у піску. Вилуплені мухи були видалені за допомогою аспіратора та перенесені в пляшки, заповнені культурним середовищем, де вони дозрівали, спарувалися та відкладали яйця, відновлюючи цикл.
Записи звуку
Нещодавно вилуплені незаймані мухи були ізольовані в 35 мл культуральних пляшках і залишені на 7-10 днів, щоб дати час для статевого дозрівання. Потім до камери запису звуку, описаної Sene & Manfrin (1998), було введено самця та самку одного й того ж роду. Для запису звуку камера з парою мух була розміщена на діафрагмі ультрачутливого мікрофона, як описано Manfrin et al. (1997). Мухи спостерігалися протягом максимум 10 хвилин або до спарювання. Усі записи були зроблені при температурі 25 ± 1ºC.
Звуковий аналіз
Електромагнітні сигнали аналогових записів були перетворені в цифрові сигнали з частотою 5 кГц за допомогою аналогово-цифрового перетворювача CAD 12/36-60 k (Lynx) та програми AqDADOS 4 (Lynx).
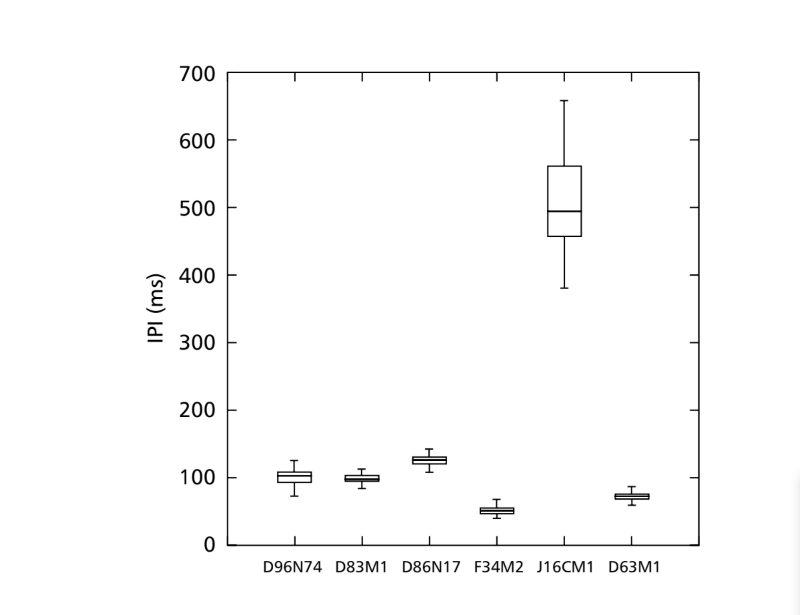
Звукові патерни шлюбного поведінки шести різних видів підгрупи fasciola описані в Таблиці 1. Також були проаналізовані звукові патерни самців D. coroica з трьох різних місць для вивчення міжпопуляційних відмінностей.
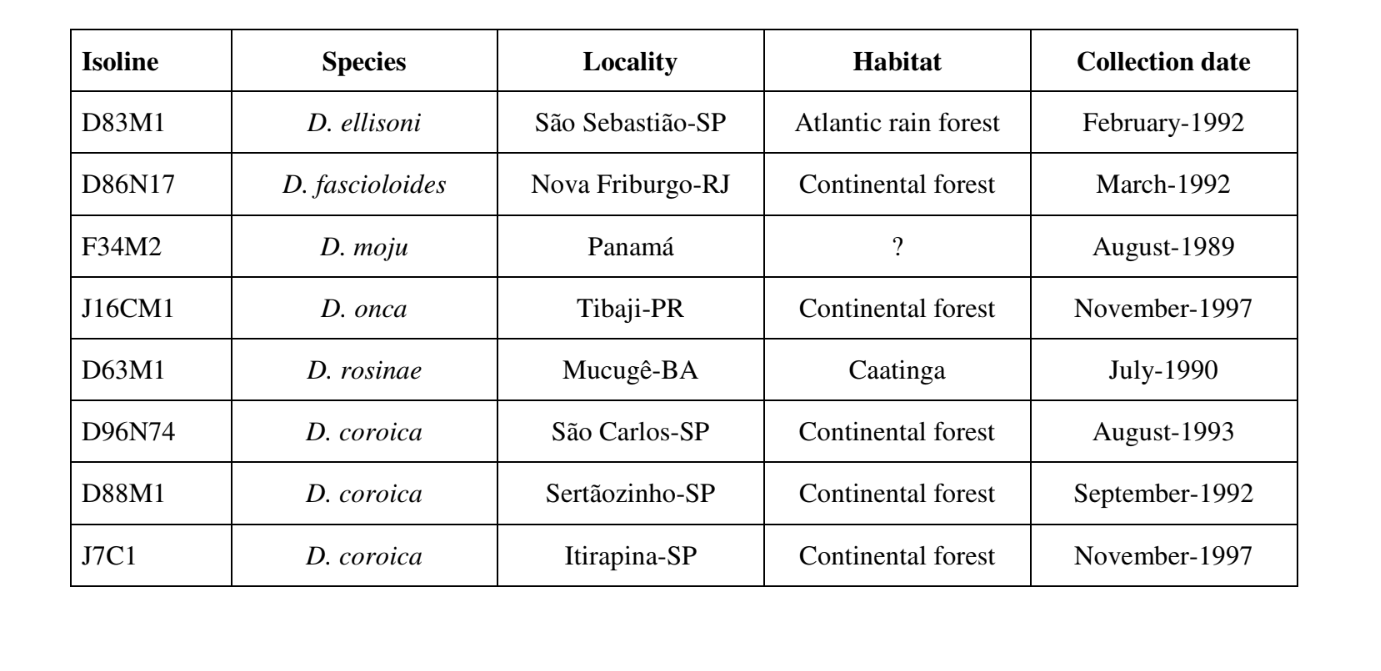
Параметри, що використовувалися для порівняльних досліджень звукових патернів, були інтервал між імпульсами (PI), інтервал між імпульсами (IPI) та частота імпульсу (IF). Значення PI та IPI були отримані безпосередньо з малюнків. Частоти IF були отримані за допомогою програмного забезпечення MATLAB (MathWorks, Inc.) з використанням швидкого перетворення Фур'є (FFT). Цей аналіз розкладає звуковий сигнал на серію хвиль (синусів і косинусів) різних частот і амплітуд. IF звуку визначається як частота хвилі з найвищою амплітудою в звуковому сигналі.
FFT може аналізувати звуки, що містять тисячі точок на мілісекунду, але максимальна ефективність аналізу досягається шляхом обмеження кількості точок до степенів двійки. З цієї причини всі інтервали, що використовувалися для аналізу IF, були обрані так, щоб мати кількість точок, рівну степеням двійки. Тому були використані інтервали по 64 точки для D. moju; 128 точок для D. rosinae та D. coroica; 256 точок для D. ellissoni; і 512 точок для D. onca та D. fascioloides. Було зроблено десять вимірювань параметрів PI, IPI та IF для кожного з 10 самців з кожної з ізоліній.
Статистичний аналіз
Лінії, описані в Таблиці 1, були проаналізовані з метою дослідження міжвидових відмінностей. Ізолінія D96N74 була обрана як представник D. coroica. Три лінії (D96N74, D88M1 та J7C1) D. coroica були протестовані окремо на предмет міжпопуляційних відмінностей. Значення параметрів інтраімпульсного інтервалу (PI), інтерімпульсного інтервалу (IPI) та інтраімпульсної частоти (IF) були проаналізовані окремо за допомогою дисперсійного аналізу (ONE-WAY ANOVA) на рівні значущості 0.05.
Результати
Рисунки 1 до 6 показують основні сонограми різних проаналізованих видів. Ізолінія D96N74 була обрана для представлення виду D. coroica. Ці сонограми та аналіз звукових імпульсів дозволяють розділити звуки courtship на три чіткі групи: 1. звуки courtship видів D. onca, D. fascioloides та D. ellisoni, в яких кожна звукова послідовність складається з послідовності коротких імпульсів, за якими слідує триль, подібний до того, що спостерігається у деяких видів підгрупи planitibia, названої Хойкала та ін. (1994) як фразова пісня. У видах D. fascioloides та D. onca послідовність імпульсів складається з чотирьох імпульсів, за якими слідує відносно довгий триль, тоді як у D. ellisoni є лише два імпульси, за якими слідує коротший триль; 2. тип, типовий для сонограми D. rosinae, з звуком courtship, що складається з простих, але подвоєних імпульсів, шаблон, також спостережений Евінгом і Міаном (1986) у деяких видах групи repleta; 3. тип, заснований на звуках courtship видів D. moju та D. coroica, з короткими імпульсами в послідовності, подібними до пульсової пісні D. melanogaster (Шилхер, 1976) та звуку A D. mercatorum (Ікеда та Маруо, 1982; Манфрін та ін., 1997).
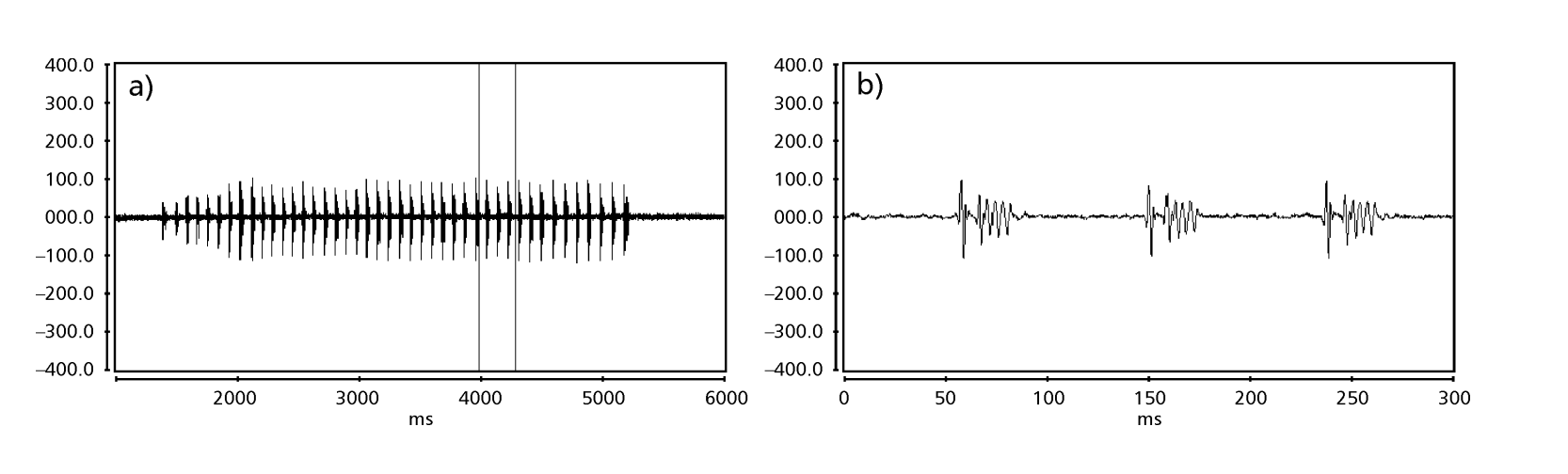
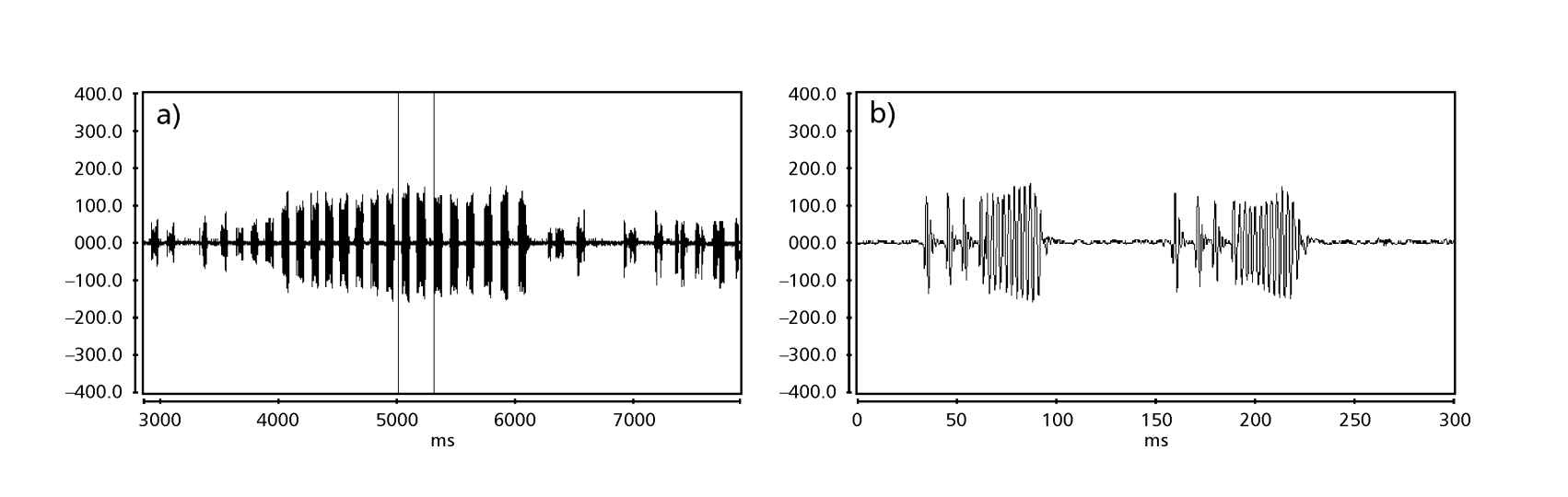
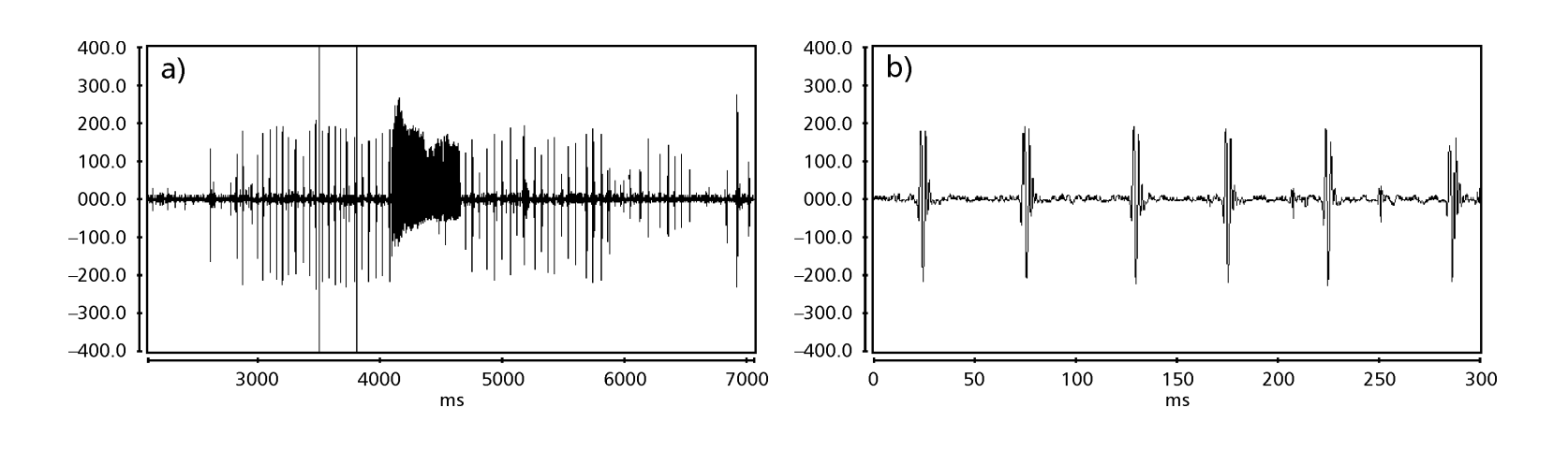
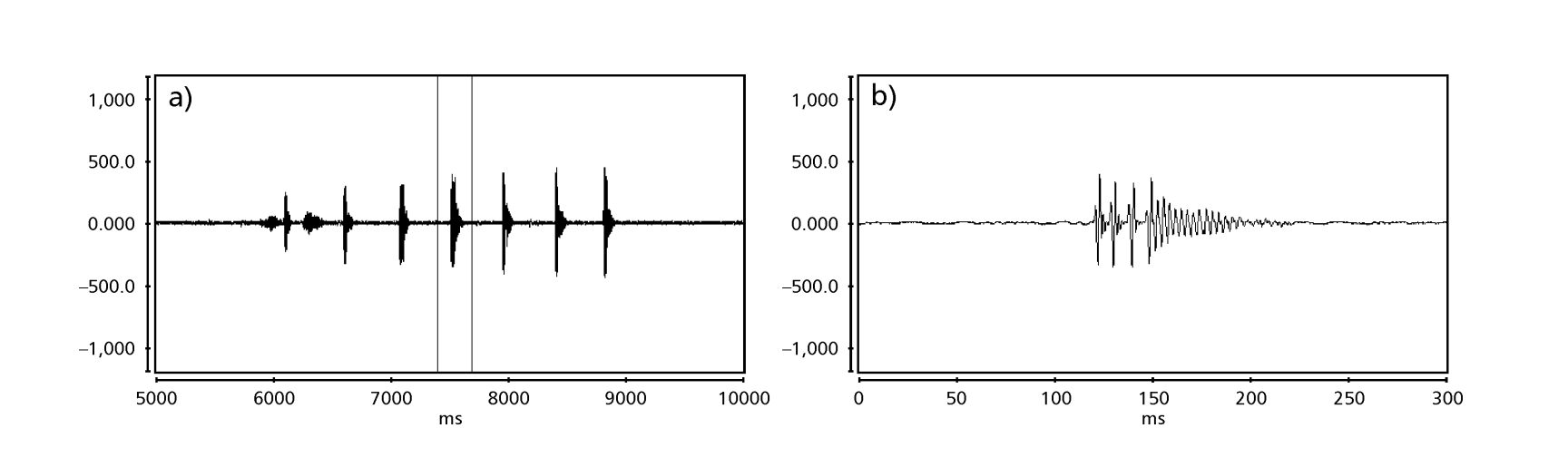
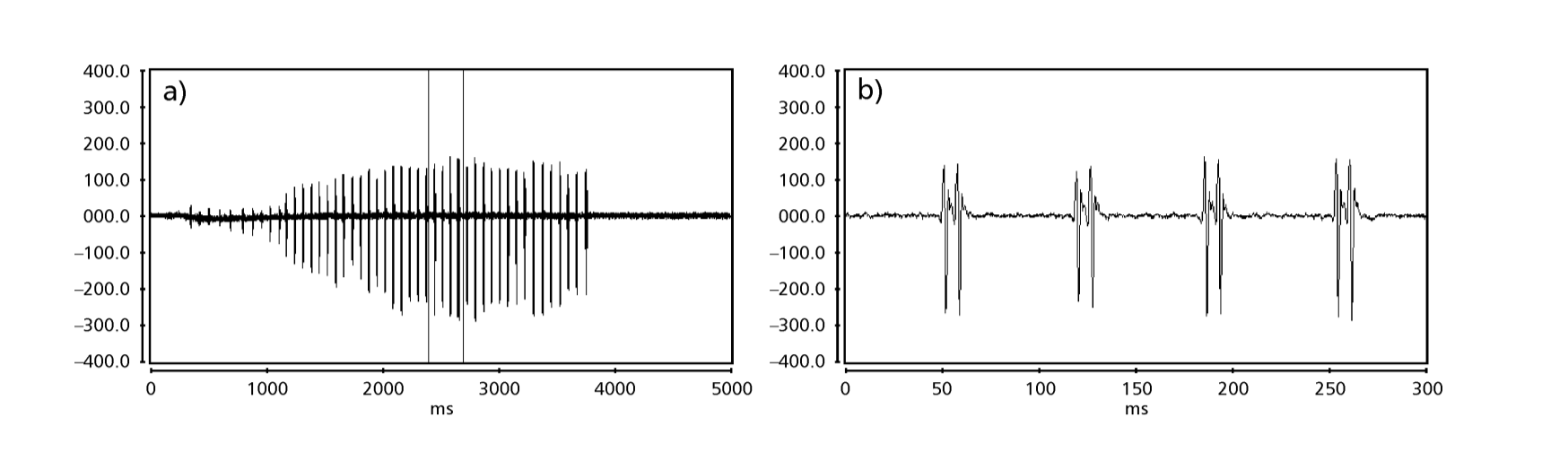
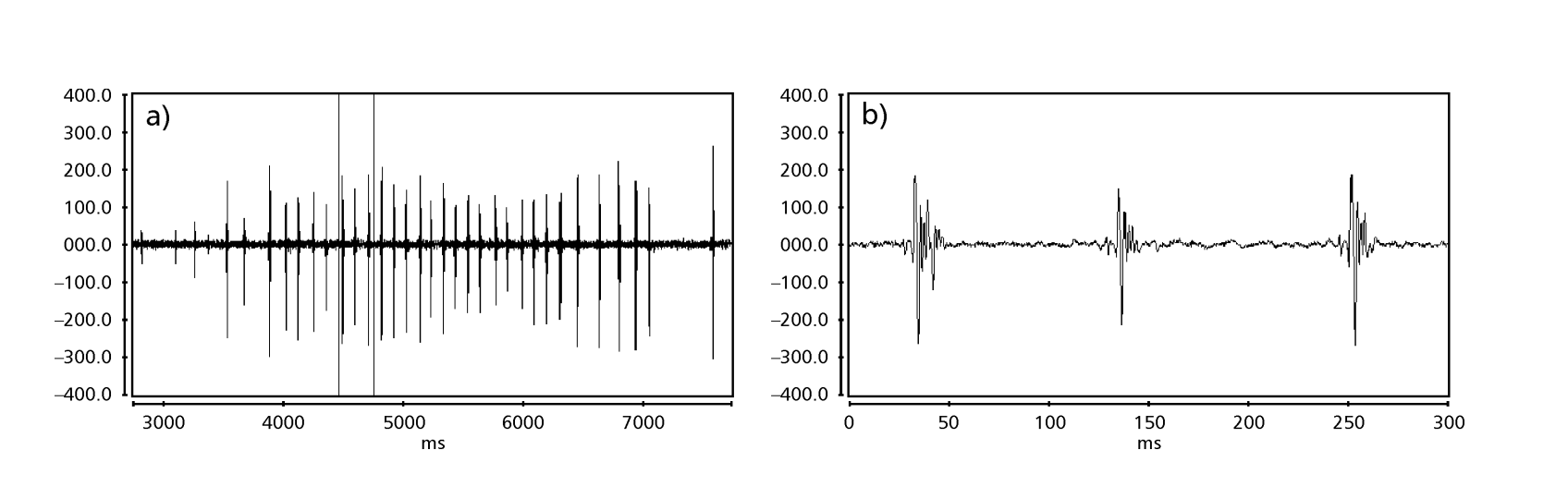
Сонограми кожного з проаналізованих видів були послідовністю одного типу звукового імпульсу. Це відкриття відрізняється від результатів досліджень, проведених на видах D. melanogaster (Schilcher, 1976), D. mercatorum (Ikeda & Maruo, 1982; Manfrin et al., 1997) та деяких видів підгрупи repleta (Ewing & Miyan, 1986), у яких були виявлені два різні типи звуків.
Таблиця 2 показує середні значення для PI (інтервал між імпульсами), IPI (інтервал між імпульсами) та IF (частота імпульсу) ізоліній, описаних у Таблиці 1. Порівняння стандартних відхилень (± sd) серед різних видів показує, що D. ellisoni та D. fascioloides мають найменшу варіацію для всіх трьох параметрів: PI (27.470 ± 2.73 та 69.859 ± 6.12), IPI (97.473 ± 5.76 та 125.757 ± 7.87) та IF (251.358 ± 19.36 та 362.901 ± 17.04), що вказує на те, що звуки шлюбного танцю цих видів є однорідними з малою внутрішньовидовою варіацією. З іншого боку, D. onca мала значну варіацію в PI (93.662 ± 31.44) та IPI (508.091 ± 73.06) і помірно високу варіацію в IF (289.260 ± 32.71) у порівнянні з іншими видами, що вказує на те, що різниці в PI та IPI, хоча й великі, не були достатніми для значного зміни форми імпульсної хвилі. Стандартні відхилення були відносно низькими для PI (7.363 ± 7.36 та 14.673 ± 1.98) та IPI (50.859 ± 5.47 та 72.340 ± 5.68) у D. moju та D. rosinae, але значення IF були відносно високими (412.501 ± 55.63 та 306.624 ± 57.80). У цьому випадку, хоча тривалість імпульсу та час від одного імпульсу до наступного були відносно однорідними в межах кожного з цих видів, форма звукового імпульсу варіювалася значно.
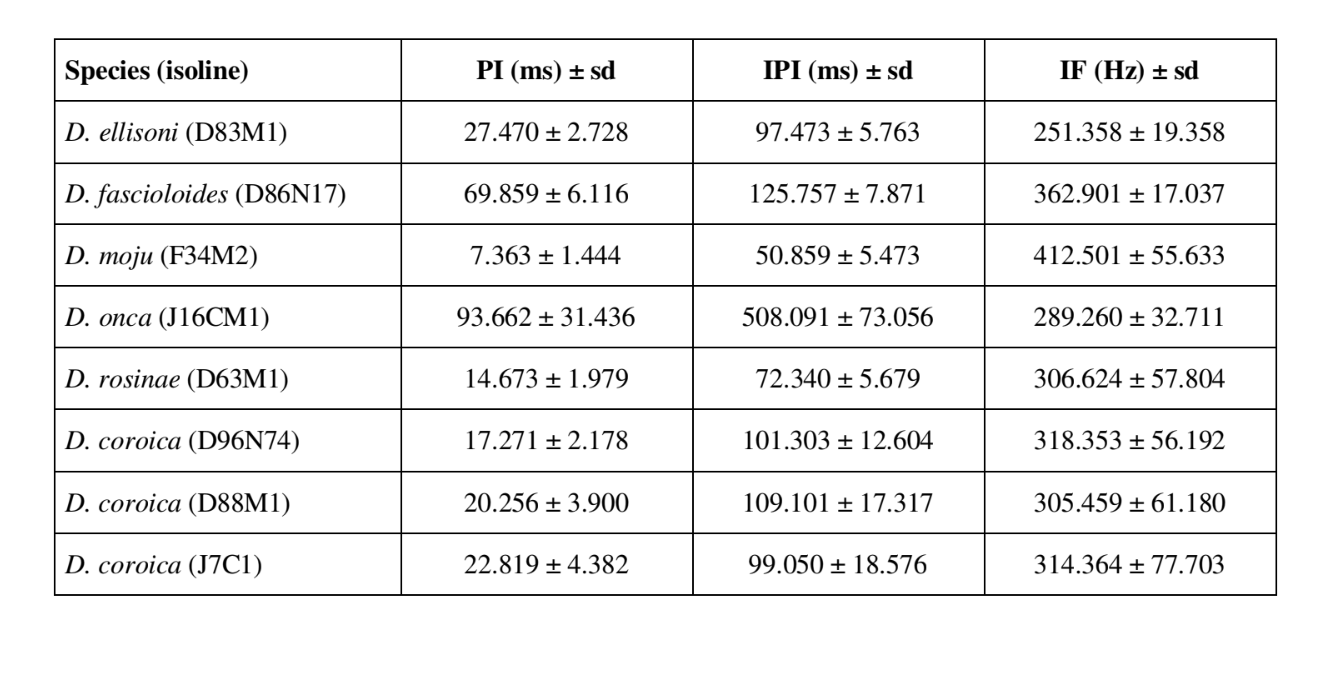
Внутрішньопопуляційна варіація в PI ліній D. coroica D96N74, D88M1 та J7C1 була відносно невеликою (17.271 ± 2.18, 20.256 ± 3.90 та 22.819 ± 4.38 відповідно), середня варіабельність в IPI (101.303 ± 12.60, 109.101 ± 17.32 та 99.050 ± 18.58) та висока варіабельність в IF (318.353 ± 56.19, 305.459 ± 61.18 та 314.364 ± 77.70), що вказує на те, що навіть при низькій внутрішньопопуляційній варіації в довжині звукового імпульсу, існувала значна варіація в формі звукового імпульсу.
Коли середні значення для параметрів PI та IPI були порівняні, різниці серед видів підгрупи fasciola можна було знайти принаймні для одного з цих параметрів. Ці різниці в звуках залицяння можна побачити на рис. 7, де значення для цих двох параметрів представлені у вигляді еліпсів, які включають 75% значень для параметрів PI та IPI. Значні різниці були виявлені в PI (ANOVA, F5,48 = 124.98, p < 0.001) та IPI (ANOVA, F5,48 = 949.35, p < 0.001) серед видів. Міжвидові різниці в IF показані на рис. 8. Значні різниці також були виявлені в значеннях IF (ANOVA, F5,48 = 23.40, p < 0.001).
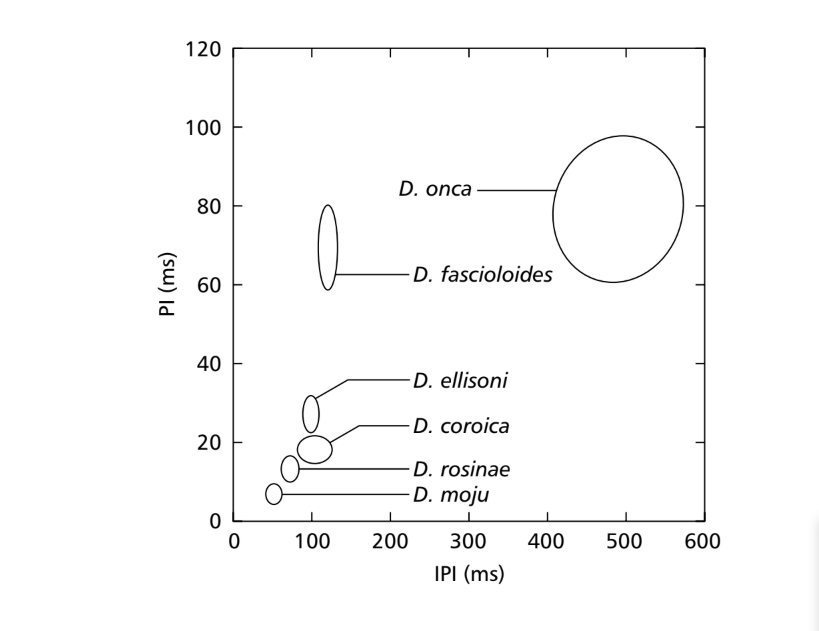
Три популяції D. coroica суттєво відрізнялися за PI (ANOVA, F2,24 = 8.75, p < 0.05), але не за IPI (ANOVA, F2,24 = 1.72, p > 0.10) та IF (ANOVA, F2,24 = 0.15, p > 0.25).
Обговорення
Хойккала та ін. (1994) у своєму аналізі звуків courtship різних видів підгрупи planitibia виявили, що звичайно можна знайти однакові типи звуків у філогенетично близьких видів. Також Хойккала & Канеширо (1993) спостерігали, що деякі тісно пов'язані види в цій підгрупі, які є симпатричними, використовують різні типи звуків у courtship.
Нур & Аквадро (1998) виявили суттєві відмінності в звуках courtship у двох тісно пов'язаних видів підгрупи obscura: D. pseudoobscura та D. persimilis. Деякі експерименти, що включали схрещування між цими двома видами, вказували на те, що самки D. pseudoobscura ймовірно підвищили свою дискримінацію проти самців D. persimilis у симпатричних популяціях. Це могло статися через відбір проти виробництва погано адаптованих гібридів, що призвело до зміни звуку courtship у цих симпатричних популяціях.
Юінг і Міян (1986), працюючи з видами групи repleta, не знайшли зв'язку між філогенією, основаною на цитологічних даних, та іншою, побудованою на основі звукових даних. Проте деякі характеристики, які мають спільними більшість вивчених видів і які також зустрічаються у видів інших груп, дозволяють нам запропонувати предковий шлюбний звук для групи repleta. Цей звук, ймовірно, складався з двох різних компонентів: початковий звук A, що складається з максимум 10 коротких і регулярних моноциклічних імпульсів, з інтервалом між імпульсами від 8 до 15 мс; і звук B, що складається з серії регулярних і моноциклічних імпульсів, але з більшою кількістю імпульсів і довшим інтервалом між імпульсами в кінці шлюбного періоду. Під час еволюції групи repleta деякі види могли втратити звук A, а інші - звук B, і у багатьох звук B став менш регулярним і більш складним.
Виходячи з цих припущень і спостерігаючи за сонограмами, отриманими для шести видів підгрупи fasciola в цьому дослідженні, ми можемо зробити висновок, що у всіх цих видів відбувалася втрата звуку A, оскільки інтервали між імпульсами завжди значно перевищують 15 мс. D. coroica має нерегулярні імпульси, з середнім інтервалом між імпульсами 101.3 мс, тоді як шлюбні звуки D. moju складаються з більш регулярних імпульсів, з середнім інтервалом між імпульсами
50.9 мс. D. rosinae має сонограму, що складається з подвоєних імпульсів, тип звуку, також виявлений Евінгом і Міаном (1986), з середнім інтервалом між імпульсами 72.3 мс. D. ellisoni, D. fascioloides та D. onca мають звуки courtship, що складаються з поліциклічних імпульсів, ймовірно, внаслідок злиття звуків A і B, з середніми інтервалами між імпульсами 97.4, 125.7 та 508.0 мс відповідно. Ці подібності в типах звуків у цих трьох видів не тісно корелюють з цитологічними даними, за винятком D. ellisoni та D. fascioloides, які були розташовані близько один до одного в філогенії на основі цитологічних ознак, вони мають два загальні злиття хромосом: 2-4F і 3-5F (Дініс, 1998). Як і у випадку з морфологічними ознаками, очікується, що тісно пов'язані види будуть ділитися деякими параметрами звуку. Проте, навіть коли філогенетичні зв'язки між видами відомі, еволюційні тенденції на основі звукових ознак ще не були добре встановлені.
Хоча звук сватання не є прикладом циркадної поведінки, дослідження пов'язали цю поведінку з експресією гена period (per), розташованого на X-хромосомі та добре охарактеризованого на молекулярному рівні. Мутації в цьому гені можуть змінювати циркадний ритм Drosophila. Генетичний аналіз, проведений Конопкою та ін. (1996), показав, що ритм звуку інтерпульсного інтервалу також підлягає впливу мутацій у гені per. Кіріакоу та Холл (1980) повідомили про ритмічну модуляцію інтерпульсного інтервалу з конкретними періодами у D. melanogaster та D. simulans відповідно 55 і 35 секунд. Ці відмінності виявилися пов'язаними з деякими варіаціями в гені per. Мутації в цьому гені, які спочатку вивчалися Конопкою та Бенцером (1971) в аналізі аномальних циркадних ритмів, викликали відповідні ефекти в ритмах інтерпульсних інтервалів звуків сватання. Мутація perShort (perS), з циркадним ритмом близько 19-20 годин, мала середній період 41.5 секунди; мутація perLong (perL), чий циркадний ритм становив 28-30 годин, мала середній період 82.1 секунди; а мутація per0 не мала ритму інтерпульсного інтервалу.
У цьому дослідженні ми виявили значні відмінності в параметрах інтраімпульсного інтервалу, інтерімпульсного інтервалу та інтраімпульсної частоти в аналізованих видах fasciola. Серед цих параметрів інтерімпульсний інтервал був тим, що відрізнявся найбільше, з малою накладкою та малою внутрішньовидовою варіацією, що свідчить про те, що цей параметр може бути частиною системи розпізнавання видів. Цей високий рівень диференціації в чоловічих звуках узгоджується з загальним консенсусом, що поведінка під час залицяння може відрізнятися більше, ніж морфологічні ознаки або інші характеристики у споріднених видах. Однак наразі неможливо знати напрямок еволюції цих різних типів звуків у підгрупі fasciola. Генетичні дослідження показали, що міжвидові відмінності в інтерімпульсних інтервалах часто зумовлені адитивними та полігенними факторами. Ці відмінності також можуть бути пояснені адаптацією до різних середовищ за відсутності генетичного потоку. Відомо, що в четвертинному періоді існували чергування циклів холодно-сухого та гарячого-вологого клімату. У циклі холодно-сухого клімату рослинність, адаптована до сухих умов, розширювалася, тоді як ліси відступали. Під час наступного циклу гарячого-вологого ситуація змінювалася (Ab’ Saber, 1977; Vanzolini, 1981). Вважається, що мухи слідували цим процесам скорочення та розширення лісів, що призвело б до тривалих періодів географічної ізоляції під час холодно-сухих циклів і до деяких областей симпатії під час гарячих-вологих циклів.
Хоча інтерпульсний інтервал може значно варіюватися серед різних видів роду Drosophila, зазвичай це не спостерігається, коли аналізуються популяції одного й того ж виду (Costa et al., 2000; Ritchie et al., 1994). Три проаналізовані популяції D. coroica мали значні відмінності лише в інтрапульсному інтервалі, без відмінностей в інтерпульсному інтервалі та інтрапульсній частоті. Можливо, що на інтерпульсний інтервал та інтрапульсну частоту діє сильний селекційний тиск. Ця низька варіабельність також може виникнути, якщо географічно відмінні популяції були генетично однорідними через постійний генетичний потік, або якщо спільний предок нещодавно колонізував ці популяції. Однак дані PI показують, що це не так.
Мало що відомо про генетичні механізми, що беруть участь у процесах видоутворення, однак спочатку необхідно зрозуміти генетику та еволюцію механізмів, які перешкоджають генетичному потоку. Неплідні та нестабільні гібриди вважаються сильними бар'єрами для генетичного потоку, але ці типи постзиготних механізмів можуть призводити до великих витрат у формі гамет та енергії. Тому відмінності в звуках courtship можуть бути важливим ізоляційним механізмом, оскільки вони обмежують або запобігають таким втратам (Welbergen et al., 1992). Аналіз цих сигналів, які є важливими для успіху спарювання, має значний інтерес для еволюційної біології, а знання архітектури звуків може допомогти довести різні моделі сексуального відбору та видоутворення.
Автори: Коста, К. Т. А. та Сене, Ф. М.
Посилання:
- Аб’ Сабер, А. Н., 1977, Простори, зайняті розширенням сухих кліматів Південної Америки, під час періодів плейстоцену. Палеоклімат, 3: 1-19.
- Чанг, Х. Ц. та Міллер, Д. Д., 1978, Звуки залицяння та спарювання у видів підгрупи Drosophila affinis. Еволюція, 32: 540-550.
- Коста, К. Т. А., Кун, Г. Ц. С. та Сене, Ф. М., 2000, Низька варіація пісні залицяння в південних та південно-східних бразильських популяціях D. meridionalis (Диптера, Drosophilidae). Rev. Brasil. Biol., 60: 53-61.
- Ковлінг, Д. Е. та Бернет, Б., 1981, Пісні залицяння та генетичний контроль акустичних характеристик у споріднених видів підгрупи Drosophila melanogaster. Анім. Поведінка, 29: 924-935.
- Дініз, Н. М., 1998, Хромосомна філогенія видів підгрупи fasciola групи repleta роду Drosophila. Дисертація на здобуття ступеня доктора, Медичний факультет Рібейран-Прету Університету Сан-Паулу.
- Евінг, А. В. та Міян, Дж. А., 1986, Статевий відбір, статева ізоляція та еволюція пісні у групі видів Drosophila repleta. Анім. Поведінка, 34: 421-429.
- Хойкала, А. та Лумме, Х., 1987, Генетична основа еволюції звуків залицяння самців групи Drosophila virilis. Еволюція, 41: 827-845.
- Хойкала, А. та Канеширо, К. Й., 1993, Зміна послідовності сигнал-відповідь, відповідальної за асиметричну ізоляцію між Drosophila planitibia та D. silvestris. Proc. Natn,. Acad. Sci., 90: 5813-5817.
- Хойкала, А. та Ісогеранен, Е., 1997, Варіація та повторюваність характеристик пісні залицяння серед диких та лабораторно вирощених самців Drosophila montana та D. littoralis (Диптера-Drosophilidae). Журнал поведінки комах, 10: 193-202.
- Хойкала, А., Лаковара, С. та Ромппайнен, Е., 1982, Поведінка при спарюванні та звуки залицяння самців групи Drosophila virilis. У: Лаковара, С. (ред.), Досягнення в генетиці, розвитку та еволюції Drosophila. Пленум, Нью-Йорк.
- Хойкала, А., Кеннет, Й. та Хой, Р. Х., 1994, Пісні залицяння видів підгрупи з малюнковими крилами Drosophila planitibia. Анім. Поведінка, 47: 1363-1374.
- Ікеда, Х. та Маруо, О., 1982, Напрямлений відбір на частоту повторення імпульсів звуку залицяння та корельовані реакції, що виникають у кількох характеристиках у Drosophila mercatorum. Японський журнал генетики, 57: 241-258.
- Коношка, Р. Дж. та Бенцер, С., 1971, Мутації годинників у Drosophila melanogaster. Proc. Natl. Acad. Sci., 68: 2112-2116.
- Коношка, Р. Дж., Кіріакоу, Ц. П. та Холл, Дж. Ц., 1996, Мозаїчний аналіз у ЦНС Drosophila циркадних ритмів пісні залицяння, що підлягають мутації годинника. Ж. Нейрогенетика, 11: 117-140.
- Кун, Г. Ц. С., Дініз, Н. М., Алвес, М. А. Р. та Сене, Ф. М., 1995, Кариотипічна конституція видів підгрупи fasciola групи repleta роду Drosophila. Rev. Bras. Gen., 18 доповнення: 282.
- Кіріакоу, Ц. П. та Холл, Дж. Ц., 1980, Мутації циркадного ритму у Drosophila melanogaster впливають на короткострокові коливання пісні залицяння самця. Proc. Natl. Acad. Sci., 77: 6729-6733.
- Манфрін, М. Х., Прадо, П. Р. Р. та Сене, Ф. М., 1997, Аналіз звукових компонентів статевого залицяння двох підвидів Drosophila mercatorum (Диптера, Drosophilidae). Rev. Brasil. Biol., 57: 349-355.
- Німс, Р. М., Духер, К., Батлін, Р. К. та Шоррокс, Б., 1997, Відмінності в пісні залицяння самців серед видів групи quinaria роду Drosophila. Журнал поведінки комах, 10: 237-246.
- Нур, М. А. та Аквадро, К. Ф., 1998, Пісні залицяння Drosophila pseudoobscura та D. persimilis: аналіз варіації. Анім. Поведінка, 56: 115-125.
- Річі, М. Г. та Глісон, Дж. М., 1995, Швидка еволюція патерну пісні залицяння у видів Drosophila willistoni. Ж. еволюційної біології, 8: 463-479.
- Річі, М. Г. та Кіріакоу, Ц. П., 1996, Штучний відбір на сигнал залицяння у Drosophila melanogaster. Анім. Поведінка, 52: 603-611.
- Річі, М. Г., Яте, В. Х. та Кіріакоу, Ц. П., 1994, Генетична варіабельність інтервалу між імпульсами пісні залицяння серед деяких європейських популяцій Drosophila melanogaster. Спадковість, 72: 459-464.
- Шільхер, Ф. В., 1976, Роль слухових стимулів у залицянні Drosophila melanogaster. Анім. Поведінка, 24: 18-26.
- Сене, Ф. М. та Манфрін, М. Х., 1998, Камера для запису пісні залицяння у Drosophila. D. I. S., 81: 203-204.
- Шорі, Х. Х., 1962, Природа звуку, що виробляється Drosophila melanogaster під час залицяння. Наука, 137: 677-678.
- Тідон-Склорз, Р. та Сене, Ф. М., 1992, Вертикальний та тимчасовий розподіл видів Drosophila (Диптера, Drosophilidae) в лісовій зоні штату Сан-Паулу, Бразилія. Rev. Brasil. Biol., 52: 311-317.
- Томару, М. та Огуму, Й., 1994, Відмінності в пісні залицяння у видів комплексу Drosophila auraria. Анім. Поведінка, 47: 133-140.
- Ванцоліні, П. Е., 1981, Квазі-історичний підхід до природної історії диференціації рептилій у тропічних географічних ізолятах. Папери Авулсос Зоол., 34: 189.
- Вілела, Ц. Р., 1983, Ревізія групи видів Drosophila repleta (Диптера, Drosophilidae). Revta. Bras. Ent., 27: 1-114.
- Вілела, Ц. Р., Перейра, М. А. Q. Р. та Сене, Ф. М., 1983, Попередні дані про географічний розподіл видів Drosophila в межах морфокліматичних доменів Бразилії. II. Група repleta. Cienc. Cult., 35: 66-70.
- Вассерман, М., 1962, Цитологічні дослідження групи repleta роду Drosophila. VI. Підгрупа fasciola. Університет Техасу Публікації., 6205: 119-134.
- Вассерман, М., 1982, Еволюція групи repleta. У: М. Ашбернер, Х. Л. Карсон та Дж. Н. Томпсон (ред.), Генетика та біологія Drosophila, т. 3b. Академічний прес, Лондон, с. 61-139.
- Вассерман, М., 1992, Цитологічна еволюція групи видів Drosophila repleta: VI. Підгрупа fasciola. У: Дослідження в генетиці II. Дослідницькі звіти про генетику Drosophila, таксономію та еволюцію. CRC Press, Inc., М. Р. Університет Техасу, Остін.
- Велберген, П., Ван Дукен, Ф. Р., Шарлоо, В. та Кьолер, В., 1992, Генетична основа статевої ізоляції між Drosophila melanogaster та D. simulans. Еволюція, 46: 1385-1398.
- Уілер, Д. А., Філдс, В. Л. та Холл, Дж. Ц., 1988, Спектральний аналіз пісень залицяння Drosophila: D. melanogaster, D. simulans та їх міжвидовий гібрид. Генетика поведінки, 18: 675-703.