Спільнота Drosophila в ксерофітних рослинностях верхньої частини басейну річки Парана-Парагвай
Машинний переклад
Оригінальна стаття написана мовою EN (посилання для прочитання) .
Анотація
Рід Drosophila є найчисленнішим у родині Drosophilidae. Деякі види є ендемічними для певних регіонів, а інші є космополітами. У Бразилії є кілька екосистем, які можна дослідити щодо складу та екологічних аспектів Drosophila. Ксерофітні території є прикладом. Вони можуть бути знайдені на півдні та південному сході Бразилії у вигляді островів, що є результатом змін палеокліматичних циклів. Метою даної роботи є надання інформації про склад спільноти Drosophila в восьми ксерофітних зонах (де росте кактус Cereus hildmaniannus ) на півдні та південному сході Бразилії. Ця робота є важливим кроком у вивченні кількісних екологічних аспектів спільноти Drosophila в ксерофітних зонах, які формують континентальні острови в неотропічному регіоні. Склад спільноти Drosophila , який було виявлено, сумісний з попередніми роботами в кількох аспектах. Екологічні індекси показали можливу позитивну асоціацію між різноманіттям і ступенем збереження досліджуваних територій. Спільноти штату Сан-Паулу продемонстрували найвищу схожість серед протестованих місць, хоча статистично значущої кореляції між індексом Жаккара та географічною відстанню не було виявлено за допомогою тесту Мантела.
Вступ
Хоча в 18 столітті Скополі, Лінней та Фабрицій були першими, хто описав види, які сьогодні відносять до родини Drosophilidae, Дуда (1924a, 1924b, 1925) може вважатися піонером у систематиці цієї групи. Більше того, Стуртевант, відомий своєю класичною працею (1921, 1939, 1942), є одним з найважливіших дослідників у цій родині. Перші дані про бразильську Drosophila спільноту були опубліковані Дудою (1925). У південно-східному регіоні Бразилії Добржанський і Паван (1943) були піонерами та описали кілька нових видів. Після 1940 року було опубліковано багато робіт про бразильську Drosophila фауну (Паван і Кунья, 1947; Добржанський і Паван, 1950; Паван, 1950; 1959), включаючи перший список видів Drosophila Бразилії (Мурао та ін., 1965). Пізніше інші автори аналізували Drosophila фауну в морфокліматичних доменах Бразилії в зусиллі краще зрозуміти географічний розподіл найбільш поширених видів (Сене та ін., 1980; Вілела та ін., 1983; Валь та ін., 1981).
Сімейство Drosophilidae складається з 65 родів та більше ніж 3,500 описаних видів, які зустрічаються в різних екосистемах по всьому світу (Bächli, 1998). Більшість родів знаходяться в тропічних регіонах. Рід Drosophila є найбільш численним і складає близько 53% від загальної кількості видів. Деякі з них є ендемічними для певних регіонів, а інші є космополітами, які в основному розповсюджені у зв'язку з людською діяльністю. Grimaldi (1990) переглянув класифікацію сімейства Drosophilidae на основі кладистичного аналізу з використанням максимальної економії. Цей автор вивчав 217 морфологічних ознак дорослих особин та 120 видів, що представляють сімейство, і дійшов висновку, що вони становлять монофілетну групу. Рід Drosophila ймовірно виник у тропіках (Stalker, 1976), і різні його члени розширили своє поширення до помірних зон, де була обрана здатність використовувати ресурси в екстремальних умовах навколишнього середовища. Цей процес, можливо, повторювався кілька разів під час еволюції роду.
Сьогодні більшість досліджень, що використовують Drosophila види, проводяться в галузях молекулярної біології та генетики, але лише на початку двадцятого століття Drosophila melanogaster була визнана корисною для генетичних аналізів (Castle, 1906). Незважаючи на велику кількість досліджень, що використовують Drosophila як модельний організм у цих галузях, бразильські види все ще потребують подальших досліджень щодо багатьох аспектів екології, систематики, генетики та еволюції.
Деякі екологічні дослідження з Drosophila були проведені, особливо щодо активності мух протягом дня (Klaczko et al., 1983; Medeiros, 2000), відповідного періоду дня для відбору зразків (Pavan et al., 1950; Belo & Oliveira-Filho, 1978) та просторового розподілу видів (горизонтальний: Dobzhanzky & Pavan, 1950; Burla et al., 1950; Pavan, 1959; Medeiros, 2000; та вертикальний: Sene et al., 1981; Kratz et al., 1982 e Tidon-Sklorz & Sene, 1992). Однак порівняння між районами отримало мало уваги (Medeiros & Klaczko, 2004, наприклад) і все ще залишається відкритою областю дослідження для бразильських Drosophila видів.
Враховуючи це, існує кілька екосистем для дослідження в Бразилії щодо складу та екологічних аспектів Drosophila. Ксерофітні області, визначені наявністю рослинності сухого типу з різними видами кактусів, є таким прикладом. Наразі, так само як і в центральних районах поширення кактусів, таких як бразильська Каа́тінга та аргентинський Чако, ці рослини можна знайти на півдні, південному сході та центральних регіонах Бразилії на островах, що є результатом циклічних палеокліматичних змін. Ці цикли чергувалися між холодними/сухими та теплими/вологими. У першому випадку ксерофітні області розширювалися з центральних районів по континенту, а в другому - вони скорочувалися, формуючи притулки (острови) між ними (Бігарелла та ін., 1975; Аб’Сабер, 1977; Ванцоліні, 1981). Отже, притулки є важливими для біорізноманіття, екологічних та еволюційних досліджень кількох груп, включаючи види Drosophila, які розмножуються специфічно в гниючій тканині кактусів, як і мухи групи D. repleta.
Метою даної роботи є надання інформації про склад спільноти Drosophila в восьми ксерофітних районах (де росте кактус Cereus hildmaniannus) на півдні та південному сході Бразилії. Це надасть цінну інформацію для подальших досліджень щодо екології, генетики та еволюції географічно ізольованих популяцій Drosophila.
Матеріали та методи
Місця та дати зборів узагальнені в Таблиці 1. Зразки були зібрані в лісах, що складаються з ксерофітної рослинності в мезофітних лісах у долинах верхньої частини басейну річки Парана-Парагвай. Ці райони перебувають на різних стадіях порушення, починаючи від використання для тваринництва поруч із дорогою (Ітатіба - SP та Сенжес - PR) до зони охорони Державного парку (Каньйон Гуартелá - PR). Усі збори були проведені в кінці вологого періоду, що відповідає лютому-травню в два різні роки (1999 та 2000).
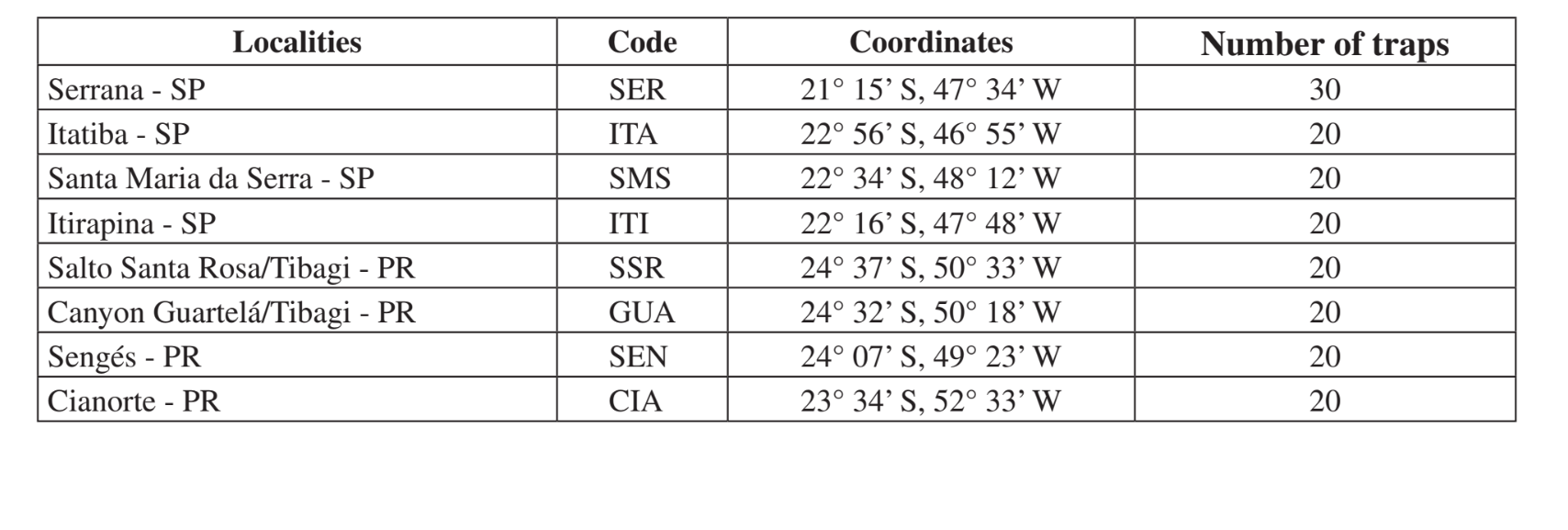
Збірки
Протягом трьох днів дорослі мухи приваблювалися до ферментованих бананів і апельсинів у відкритих пастках, розташованих не ближче ніж 15 м одна від одної, підвішених на гілках на висоті ± 1,5 м від землі. Після цього періоду мухи були спіймані ентомологічними сітками, поміщені в скляні банки з культурним середовищем і доставлені в лабораторію живими, де їх ідентифікували.
Важко ідентифікувати представників груп Drosophila repleta та D. willistoni до певного ступеня. Самки першої групи вважаються невизначеними, якщо діагностичною характеристикою є адеагус самця. Щодо групи Drosophila willistoni, ідентифікація за генетичними ознаками є більш надійною, але не проводилася. Тому криптичні види були згруповані як “repleta група” та “D. willistoni група”, відповідно. Невизначені види “repleta групи” не враховувалися в аналізах.
Аналіз даних
Для якісної перевірки появи видів був використаний Метод Постійності Появи (Dajoz, 1983). Значення постійності (c) отримували шляхом ділення кількості колекцій, в яких з'являвся один вид, на загальну кількість колекцій, а потім множення цього результату на 100. Види з індексом c ≥ 50 вважалися постійними. Допоміжними видами були ті, у яких 25 ≤ c < 50. Випадкові види мали c < 25. Види, що з'являлися лише в одній місцевості, вважалися ексклюзивними.
Drosophila спільноти аналізувалися за допомогою кількох екологічних індексів: Бергера-Паркера (1/d), Шеннона-Вінера (H’), Симпсона (D), Маргалефа (DMg), Менгініка (DMn) та Жаккара (J). Для визначення, чи існують відмінності в медіанній чисельності між спільнотами, використовували непараметричний ANOVA, Крускала-Уолліса. Після цього було проведено множинне порівняння між зразками (аналогічно тесту Тьюкі), щоб визначити, чи відбулися значні відмінності між зразками (Zar, 1999).
Для перевірки взаємозв'язків між спільнотами було проведено два кластерних аналізи:
1) використовуючи присутність та відсутність видів; та 2) використовуючи кількість зразків, зібраних для кожного виду в кожній області. Фенограми були побудовані в програмному забезпеченні Minitab® для Windows Release 10.1, вибираючи методи евклідової відстані та одиничного зв'язку в обох підходах.
Щоб проаналізувати, чи існує асоціація між подібностями спільнот і географічними відстанями, був проведений тест Мантела за допомогою програмного забезпечення TFPGA (Miller, 1997), порівнюючи дві матриці: індекси Жаккара та географічні відстані між парами спільнот.
Результати та обговорення
У цьому дослідженні ми змогли ідентифікувати загалом 10,464 мухи, що належать до 4 підродів, 10 груп і принаймні 30 різних видів Drosophila (Таблиця 2). Постійні види (c ≥ 50) становили приблизно 63% від загальної кількості зібраних видів (19 з 30). Сім видів вважалися допоміжними (~ 23%), а 4 випадкові види були знайдені (~ 13%). Усі підроди показали постійні види, за винятком підроду Dorsilopha , який показав лише один випадковий вид (Drosophila busckii).
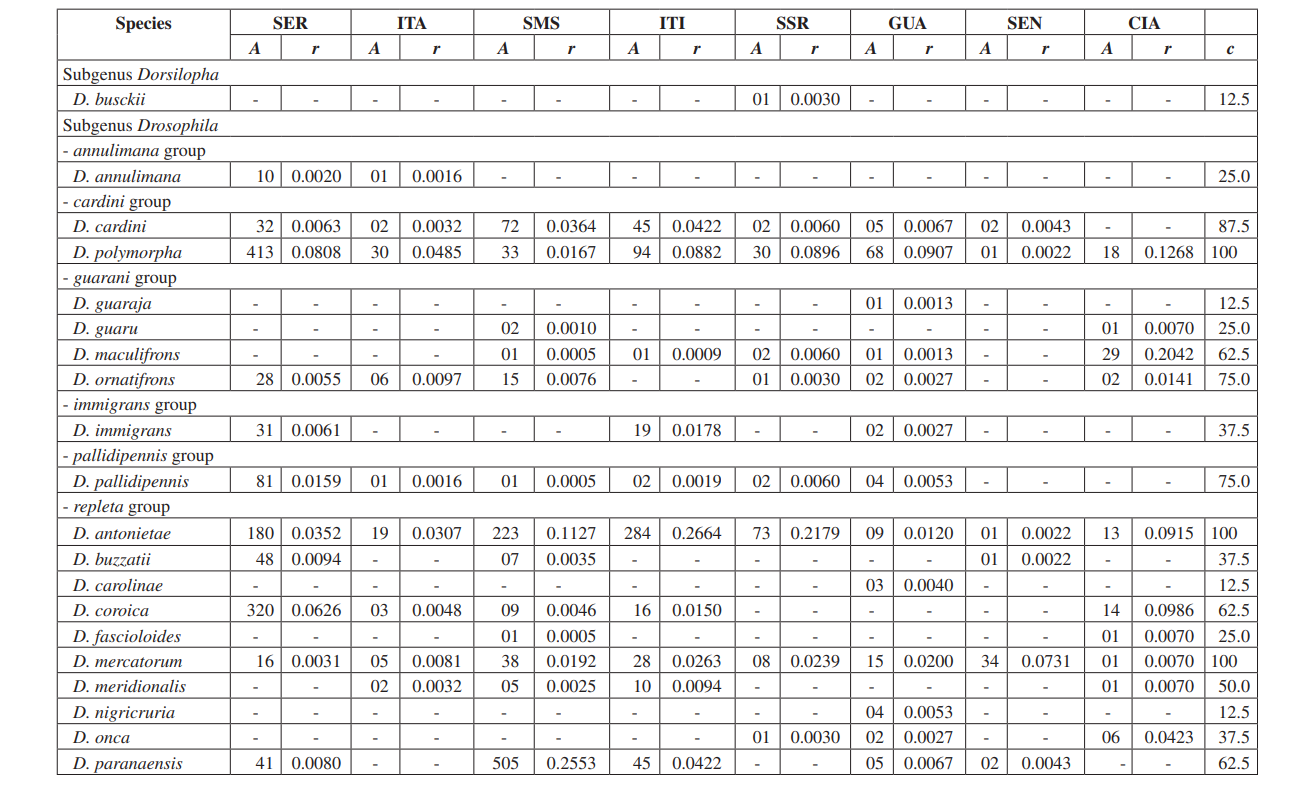
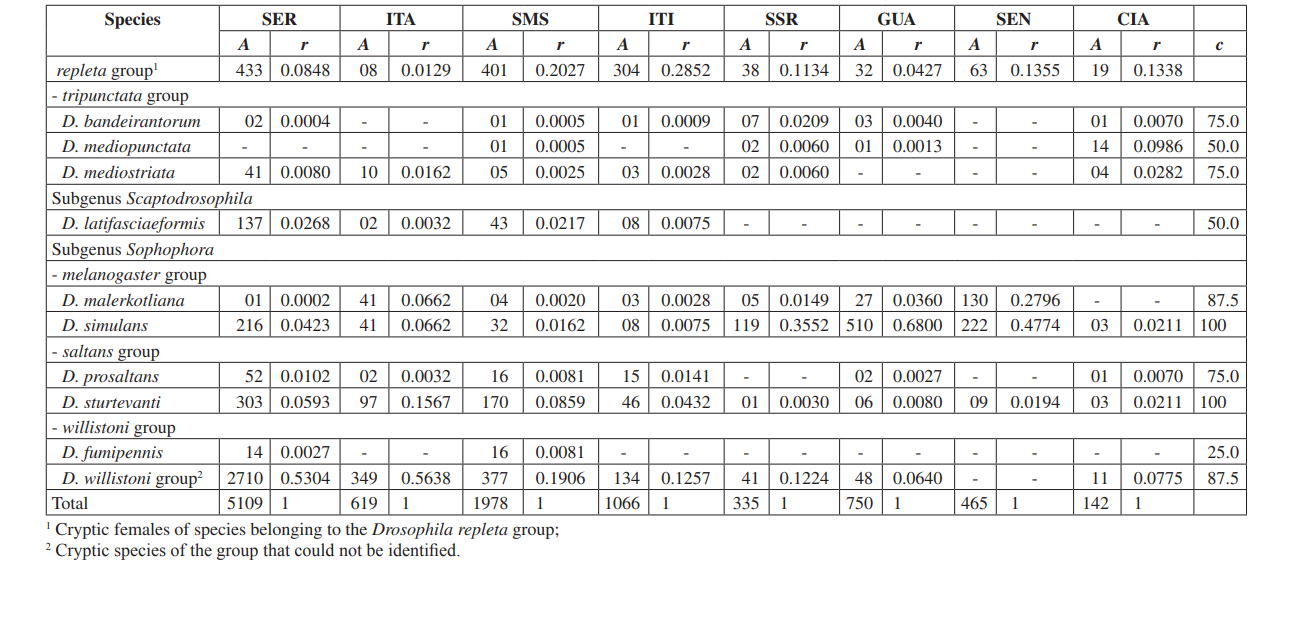
П'ять видів (Drosophila polymorpha, D. antonietae, D. mercatorum, D. simulans та D. sturtevanti) вважалися численними і були зібрані у всіх 8 районах (Серрана - SP, Ітатіба - SP, Санта-Марія-да-Серра - SP, Ітірапіна - SP, Сальто-Санта-Роза - PR, Каньйон Гуартелá - PR, Сенжес - PR та Сіанорт - PR). Цей результат узгоджується з попередніми обстеженнями Drosophila.
Drosophila polymorpha була зібрана від Гватемали до Бразилії та в різних типах середовищ: ліси, серрадо, рестінга, а також асоційована з присутністю людини. Це відносно численний вид у різних морфокліматичних доменах, за винятком каатінги (Сене та ін., 1980). Drosophila antonietae очікувалося, що буде зібрана у всіх районах, оскільки це кактусофільний вид, який зустрічається на півдні та південному сході Бразилії, а також на півночі східного кордону аргентинського Чако. Цей вид завжди зустрічається в асоціації з колонним кактусом Cereus hildmaniannus у ксерофітній рослинності галерейних і мезофітних лісів у долинах басейну річки Парана-Уругвай (Тідон-Склорз & Сене, 2001). Цей шаблон розподілу, який ізолює Drosophila antonietae від інших видів кластера D. buzzatii, збігається з визначеними шаблонами інших двокрилих, а також з регіонами ендемізму, що свідчить про вікаріантні події, такі як зоогеографічні бар'єри, встановлені в більш ранніх геологічних епохах або кліматичні події, які перешкоджали генетичному потоку, що дозволило накопичувати відмінності між популяціями (Аморін & Піреш, 1996). Цей шаблон розподілу також спостерігається щодо розподілу рослин, обмеженого до ксерофітних регіонів каатінги та чако (Дювернель & Інес, 2000).
Drosophila mercatorum є досить поширеним у природних середовищах Південної Америки, особливо в відкритих місцях (Sene та ін., 1981; Vilela та ін., 1983).
Drosophila simulans є інтродукованим видом, який був зібраний у великій кількості в кількох місцях (Dobzhansky & Pavan, 1950; Pavan, 1959; Sene та ін., 1980; Tidon-Sklorz & Sene, 1992), також в відкритих місцях (Perondini та ін., 1979).
Drosophila sturtevanti є широко розповсюдженим видом у Центральній та Південній Америці, що зустрічається в різних морфокліматичних доменах (Tidon-Sklorz & Sene, 1999).
Ексклюзивні види з'явилися в Salto Santa Rosa - PR (Drosophila busckii) та Canyon Guartelá - PR (Drosophila guaraja, D. carolinae та D. nigricruria). Drosophila busckii не очікувалося знайти в Salto Santa Rosa - PR, оскільки це інтродукований вид у Бразилії, який часто асоціюється з середовищами, зміненими людиною (Tidon-Sklorz & Sene, 1999). Однак його було зібрано в природних середовищах, переважно в сухих відкритих рослинностях (Bizzo & Sene, 1982; Tidon-Sklorz & Sene, 1992). Серед ексклюзивних видів, зібраних у Canyon Guartelá - PR, Drosophila guaraja був зареєстрований у Бразилії лише в Boracéia - SP (Val & Kaneshiro, 1988). Отже, це перший запис про нього поза цією місцевістю в Бразилії. Drosophila carolinae було зібрано лише в тропічних лісах Бразилії (Vilela, 1983), і з цієї причини не очікувалося його виявлення в зонах ксерофітної рослинності. Однак Canyon Guartelá - PR є однією з найзбереженіших територій, що досліджуються, а кактуси розподілені під лісовою зоною в межах Державного парку, що може пояснити його присутність. Drosophila nigricruria є єдиним ексклюзивним видом, який має широке географічне поширення. Проте його було зібрано лише в цій області в цьому дослідженні. Medeiros & Klaczko (2004) також зібрали цей вид на фермі Barreiro Rico (між 48° 05’ 11’’ W – 22° 41’ 15’’ S та 48° 04’ 52’’ W – 22° 41’ 26’’ S), на внутрішньому плато, яке має напівлистяний ліс і є одним із трьох залишкових лісів у штаті Сан-Паулу, де ці автори проводили збори.
Підрід Drosophila показав найвищу різноманітність (7 груп і 22 види), з 7 видами, що належать до групи Drosophila repleta . Друга за величиною різноманітність була виявлена в підроді Sophophora (3 групи і 6 видів). Це розподіл різноманітності серед підродів спостерігався в інших дослідженнях (Sene та ін., 1981; Tidon-Sklorz & Sene, 1995; Tidon-Sklorz & Sene, 1999; Medeiros & Klaczko, 2004).
Введений вид дрозофіл, Zaprionus indianus (Vilela, 1999), був знайдений у всіх колекціях (1,459 зразків у Серрані; 317 в Ітатібі; 409 у Санта-Марія-да-Серра; 302 в Ітірапіні; 42 в Сальто-Санта-Роза; 117 в Гуартелі; 598 в Сенгесі; і 8 в Сіанорті). Події, пов'язані з біологічними інвазіями, поділяються на три категорії: прибуття, встановлення та інтеграція (Vermeij, 1996). Згідно з Moraes та ін. (2000), Zaprionus indianus пройшов через всі три етапи і, ймовірно, конкурує невигідно з іншими подібними видами на південному сході Бразилії, оскільки спостерігалося падіння відносної частоти. Введення екзотичного виду Drosophila в Бразилії не є рідкісною подією. У 1976 році Вал & Сене (1980) зібрали велику кількість Drosophila malerkotliana на північному сході Бразилії, вид, який не був зафіксований у великих попередніх колекціях. Наразі в наших опитуваннях Drosophila malerkotliana збирається майже по всьому Бразилії, хоча завжди з низькою відносною частотою.
Двадцять три різні види були зібрані в Санта-Марія да Серра - SP, 20 видів у Серрані - SP та в Каньйоні Гуартела - PR, 18 в Ітірапіні - SP, 17 у Сіанорті - PR, 16 в Ітатібі - SP та Сальто Санта Роса - PR і 9 у Сенжесі - PR (Таблиця 3). Медеїрос і Клацко (2004) описали останній інвентаризацію фауни Drosophila в Бразилії, проведену в трьох лісах, що залишилися в штаті Сан-Паулу. Ці автори отримали 57, 76 і 90 різних видів у трьох вивчених районах. Ці числа видів більші, ніж спостережені тут, можливо, внаслідок відмінностей у методі збору або просто тому, що опитані райони відрізняються за своєю рослинною структурою (лісова та ксерофітна рослинність). Найвища загальна чисельність була виявлена в Серрані - SP (4676 екземплярів), а найнижча в Сіанорті - PR (123 екземпляри) (Таблиця 3).
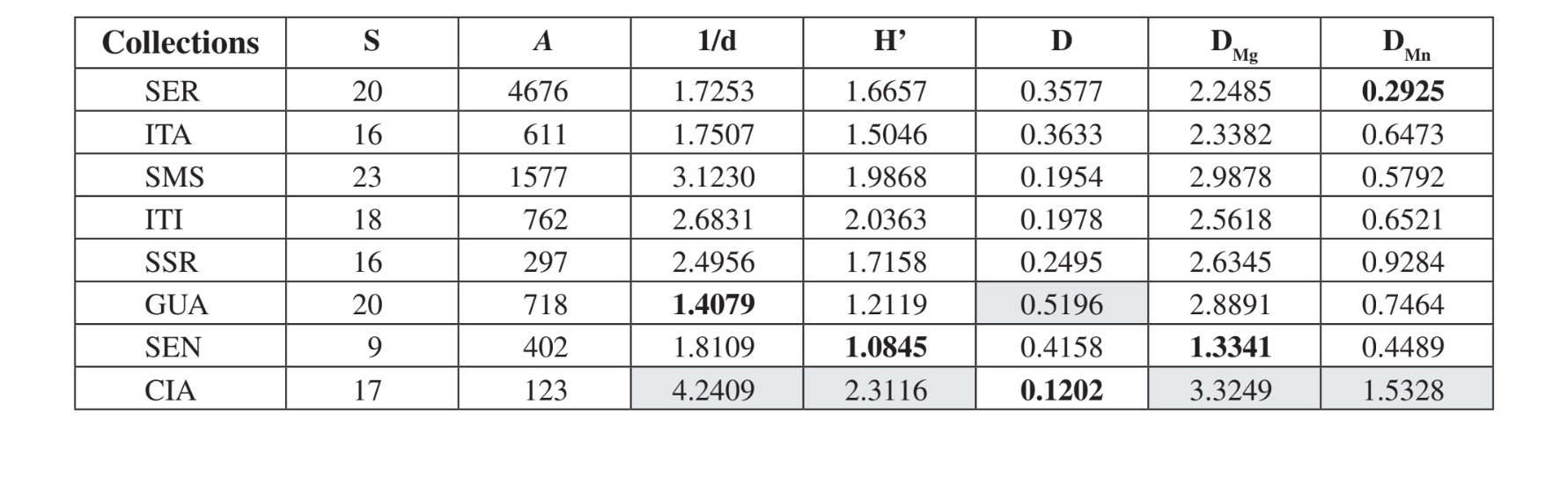
Згідно з екологічними індексами (Таблиця 3), Cianorte - PR продемонстрував вищі значення майже за всіма індексами (1/d = 4.2409; H’ = 2.3116; DMg = 3.3249; DMn = 1.5328), за винятком Симпсона (D), який показав, що Canyon Guartelá - PR є найбільш
різноманітною територією (D = 0.5196). Canyon Guartelá - PR також показав один з найвищих індексів у ще двох випадках (DMg = 2.8891; DMn = 0.7464). Sengés - PR, Itatiba - SP та Serrana - SP показали найнижчі значення за більшістю індексів. Santa Maria da Serra - SP, Itirapina - SP та Salto Santa Rosa - PR показали проміжні значення за більшістю індексів. Ці спостереження узгоджуються з поверхневою оцінкою деградації в цих районах. Cianorte - PR розташований поруч з дорогою, але на високому схилі річки Ligeiro, з важким доступом для людської деградації. Canyon Guartelá - PR є лісовою територією всередині державного парку Canyon Guartelá штату Парана, як вже згадувалося раніше. Тому це є високо збережена територія. Sengés - PR є найбільш деградованою територією, поруч з дорогою, де займаються тваринництвом і під високим впливом людської діяльності. Serrana - SP є збереженою територією на вершині пагорба, до якої важко дістатися. Нижчі індекси різноманітності, знайдені для цієї території, повинні бути пов'язані з більшою кількістю пасток, використаних у цьому зборі (Таблиця 1). Інші території, як очікувалося, мали б проміжні значення різноманітності, оскільки це невеликі охоронювані території всередині ферм.
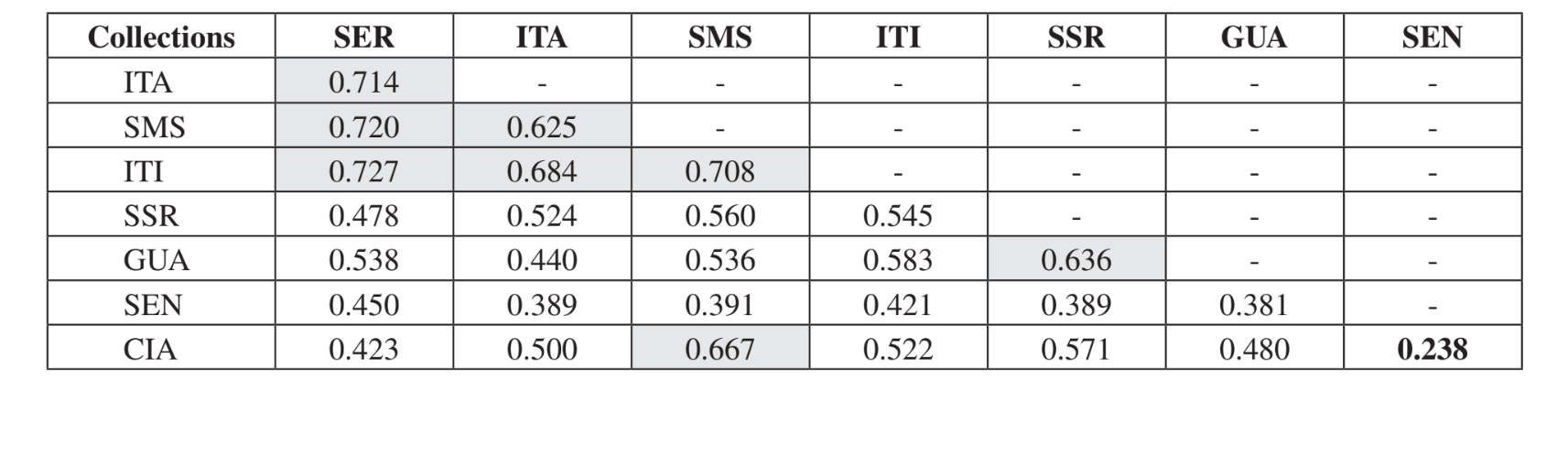
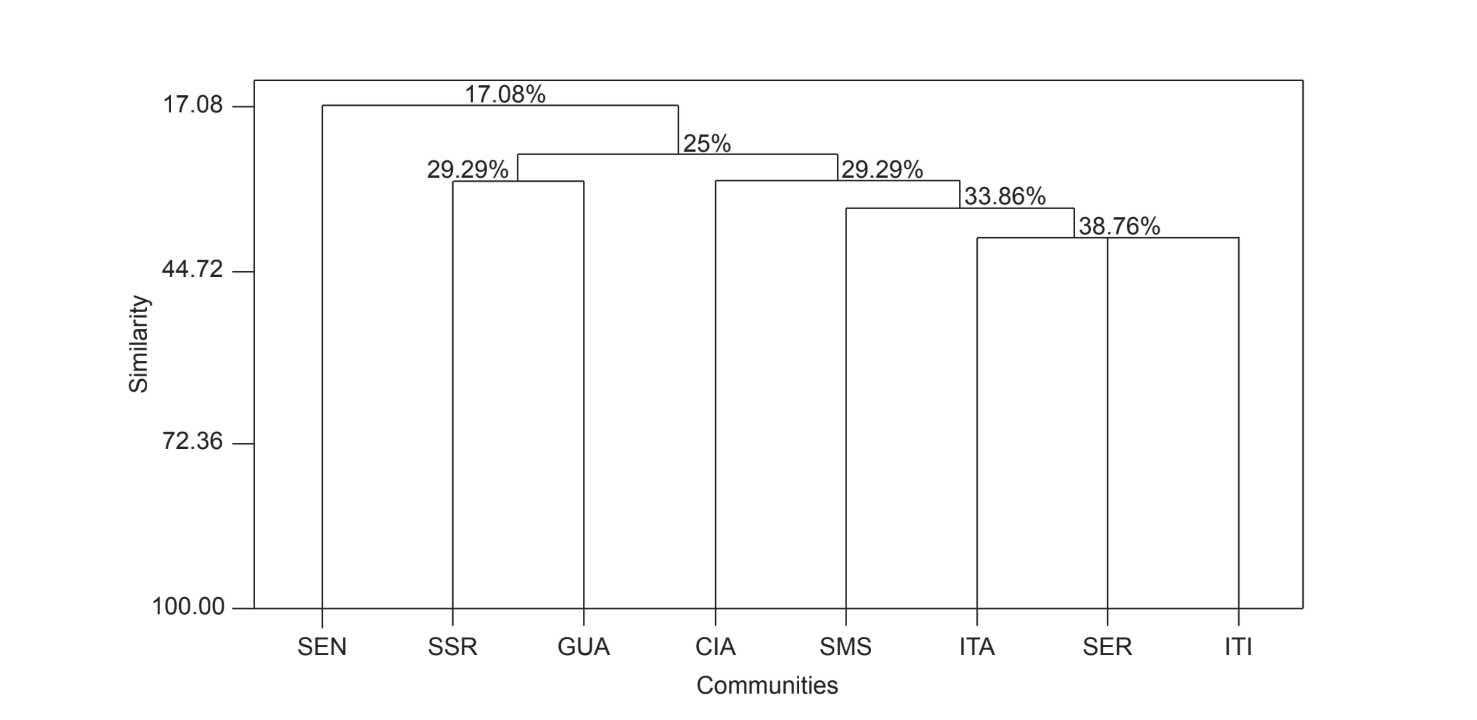
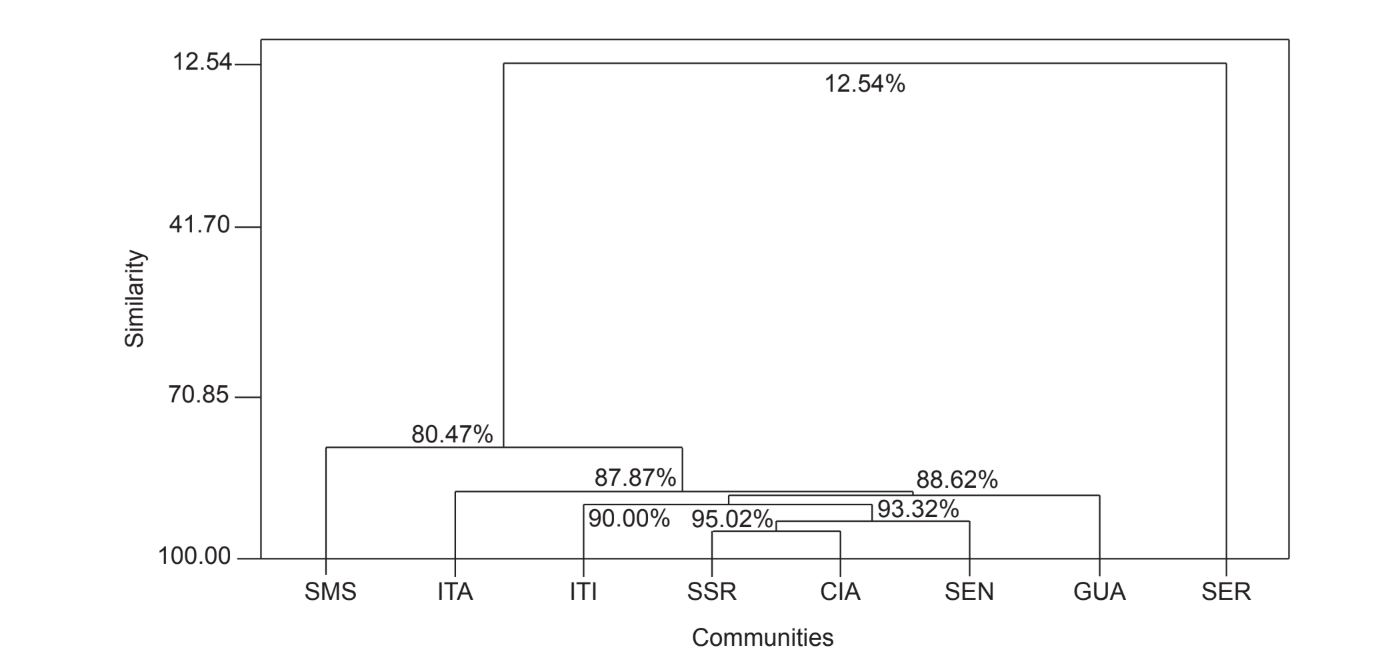
Парне порівняння спільнот за допомогою індексу Жаккара (Таблиця 4) показало, що спільноти штату Сан-Паулу (Серрана - SP, Ітатіба - SP, Санта-Марія да Серра - SP та Ітірапіна - SP) мали найвищу схожість серед усіх порівнянь (J > 0.62). Того ж патерну не спостерігали серед спільнот штату Парана, а також між обома штатами, за винятком індексу, отриманого між Санта-Марія да Серра - SP та Сіанорт - PR (J = 0.667), і між Сальто Санта Роза - PR та Каньйон Гуартелá - PR (J = 0.636). Кластерний аналіз за даними присутності та відсутності видів (Рис. 1), незважаючи на низький рівень схожості (33.86%), також об'єднав усі спільноти штату Сан-Паулу. Найбільш відмінною спільнотою була Сенгес - PR. Рис. 2 ілюструє кластерне групування всіх спільнот за даними абсолютної чисельності (A). Було виявлено вищі рівні схожості, але чіткої асоціації спільнот не було знайдено. На цій фігурі Серрана - SP була найвідміннішою спільнотою.
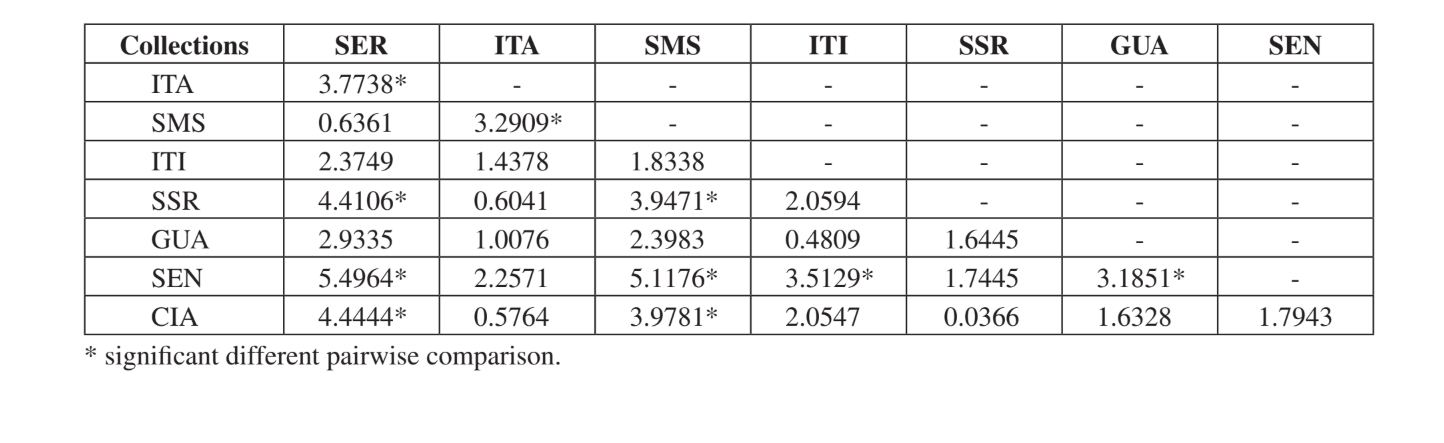
Тест Крускала-Уолліса, скоригований для зв'язків, показав, що громади суттєво відрізняються, коли було проведено загальне порівняння (H = 24.24; d.f. = 7; p = 0.001). Множинні парні порівняння громад представлені в Таблиці 5. Громади Серрана - SP, Санта-Марія да Серра – SP та Сенгес - PR були найбільш відмінними, коли їх порівнювали індивідуально з іншими (4 суттєво різних порівняння для кожної). Ітірапіна - SP та Каньйон Гуартелá - PR були найбільш схожими громадами, коли їх порівнювали з кожною вибраною громадою (лише одна суттєва різниця для кожної). Усі ці порівняння (індекс Жаккара, кластерний аналіз та множинні парні порівняння) показали, що вищий рівень схожості спостерігався серед громад штату Сан-Паулу, хоча позитивної суттєвої кореляції не було виявлено в тесті Мантела, що порівнює індекс Жаккара та географічні відстані (r = – 0.3235; p = 0.0839).
Опитування складу спільноти є важливим етапом для кількох досліджень у широкому спектрі областей, таких як генетика, екологія та еволюція. Ця робота додає нові знання щодо складу та різноманіття спільноти Drosophila в ксерофітних районах, які формують континентальні острови в неотропічному регіоні. Цей підхід важливий для подальших досліджень у будь-якій галузі, що розглядає Drosophila як модель.
Автори: Mateus, R. P., Buschini, M. L. T. та Sene, F. M
Посилання:
- Ab’saber, A. N., 1977, Простори, зайняті розширенням сухих кліматів Південної Америки, під час періодів плейстоцену. Paleoclimas, 3: 1-19.
- Amorin, D. S. & Pires, M. R. S., 1996, Неотропічна біогеографія та метод для максимального оцінювання біорізноманіття. In: C. E. M. Bicudo & N. A. Menezes (eds.), Біорізноманіття в Бразилії, перший підхід, CNPq, Сан-Паулу.
- Bächli, G., 1998, Сімейство Drosophilidae. In: L. Papp & B. Darvas (eds.), Внески до посібника з палеарктичних двокрилих. III. Вищі брахіцери, Science Herald, Будапешт.
- Belo, M. & Oliveira-Filho, J. J., 1978, Домашні види Drosophila. V. Впливи екологічних факторів на кількість захоплених особин. Rev. Bras. Biol., 36: 903-909.
- Bigarella, J. J., Andrade-Lima, D. & Riehs, P. J., 1975, Зауваження щодо палеокліматичних змін у розподілі деяких видів рослин і тварин у Бразилії. An. Acad. Brasil. Ciênc., 41: 411-464.
- Bizzo, N. M. V. & Sene, F. M., 1982, Дослідження природних популяцій Drosophila з Перуїбе (SP), Бразилія (Diptera, Drosophilidae). Rev. Bras. Biol., 42: 539-544.
- Burla, H., da Cunha, A. B., Cavalcanti, A. G. L., Pavan, C. & Dobzhansky, T., 1950, Щільність популяції та швидкість розсіювання в бразильській Drosophila willistoni. Ecology, 31: 393-404.
- Castle, W. E., 1906, Інбридинг, схрещування та стерильність у Drosophila. Science, 23: 153.
- Dajoz, R., 1983, Екологія загальна. 471p. Видавництво Vozes, Петрополіс.
- Dobzhansky, T. & Pavan, C., 1943, Дослідження бразильських видів Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 36 (Biol. Geral, 4): 1-72.
- Dobzhansky, T. & Pavan, C., 1950, Місцеві та сезонні варіації в відносних частотах видів Drosophila у Бразилії. J. Anim. Ecol., 19: 1-14.
- Duda, O., 1924a, Внесок у систематику Drosophilidae з особливою увагою до палеарктичних та орієнтальних видів (Diptera). Arch. Naturgesch., 90: 172- 234.
- Duda, O., 1924b, Ревізія європейських видів роду Drosophila Fallén (Diptera). Ent. Medd., 14: 246-313.
- Duda, O., 1925, Костариканські Drosophilidae угорського Національного музею в Будапешті. Annls hist.- nat. Mus. natn. hung., 22: 149-229.
- Duvernell, D. D. & Eanes, W. F., 2000, Контрастуюча молекулярна популяційна генетика чотирьох гексокіназ у Drosophila melanogaster, D. simulans та D. yakuba. Genetics, 156: 1191-1201.
- Grimaldi, D. A., 1990, Філогенетична переглянута класифікація родів у Drosophilidae (Diptera). Bull. Am. Mus. nat. Hist., 197: 1-139.
- Klaczko, L. B., Powell, J. R. & Taylor, C. E., 1983, Drosophila приманки та дріжджі: види, що приваблюються. Oecologia, 59: 411-413.
- Kratz, F. L., Pinto, L. G., Brandão, D. & Faria, L. G., 1982, Висота польоту та просторовий розподіл у Drosophila. Ciênc. Cult., 34: 203-209.
- Medeiros, H. F., 2000, Асамблеї видів Drosophila (Diptera; Drosophilidae) та впливи курсів води на їх розподіл у двох лісах Сан-Паулу, 98p. Дисертація (Магістратура) – Інститут біології IB/UNICAMP, Кампінс.
- Medeiros, H. F. & klaczko, L. B., 2004, Скільки видів Drosophila (Diptera, Drosophilidae) залишилося описати в лісах Сан-Паулу, Бразилія? Списки видів трьох лісових залишків. Biota Neotropica, 4: 1-12.
- Miller, M. P., 1997, Інструменти для аналізу популяційної генетики - TFPGA - 1.3: Програма для Windows для аналізу алозимних та молекулярних даних популяційної генетики.Комп'ютерне програмне забезпечення, розповсюджене автором.
- Moraes, E. M., Mateus, R. P. & Sene, F. M., 2000, Виникнення та коливання в розмірі популяції нещодавно введеного виду Drosophilidae в Бразилії. In: АБСТРАКТИ XXI МІЖНАРОДНОГО КОНГРЕСУ ЕНТОМОЛОГІЇ, КНИГА I. Proceedings..., 271p.
- Mourão, C. A., Gallo, A. J. & Bicudo, H. E. M. C., 1965, Про систематику Drosophila в Бразилії, з описом D. mendeli sp.n. та “Список бразильських видів роду Drosophila”. Ciênc. Cult., 17: 577- 585.
- Pavan, C., 1950, Бразильські види Drosophila. II. Bolm. Fac. Filos. Ciênc. S. Paulo, 111 (Biol. Geral, 8): 1-37.
- Pavan, C., 1959, Взаємозв'язки між природними популяціями Drosophila та навколишнім середовищем. Bolm. Fac. Filos. Ciênc. S. Paulo, 221 (Biol. Geral, 11): 1-81.
- Pavan, C. & da Cunha, A. B., 1947, Бразильські види Drosophila. Bolm. Fac. Filos. Ciênc. S. Paulo, 86 (Biol. Geral, 7): 3-47.
- Pavan, C., Dobzhansky, T. & Burla, H., 1950, Діурнальна поведінка деяких неотропічних видів Drosophila. Ecology, 31: 36-43.
- Perondini, A. L. P., Sene, F. M. & Mori, L., 1979, Патерн та поліморфізм деяких Drosophila simulans естераз у Бразилії. Egypt. J. Genet. Cytol., 8: 263-268.
- Sene, F. M., Val, F. C., Vilela, C. R. & Pereira, M. A. Q. R., 1980, Попередні дані про географічний розподіл видів Drosophila в межах морфокліматичних доменів Бразилії. Pap. Avul. Dep. Zool. Sec. Agric., 33: 315-326.
- Sene, F. M., Pereira, M. A. Q. R., Vilela, C. R. & Bizzo, N. M. V., 1981, Вплив різних способів встановлення приманок для збору мух Drosophila в трьох природних середовищах. D. I. S., 56: 118-121..
- Stalker, H. D., 1976, Дослідження хромосом у диких популяціях D. melanogaster. Genetics, 82: 323-347.
- Sturtevant, A. H., 1921, Північноамериканські види Drosophila. Carnegie Inst. Wash. Publ., 301: 1-150.
- Sturtevant, A. H., 1939, Про підрозділ роду Drosophila. Proc. Natl. Acad. Sci. USA, 3: 137-141.
- Sturtevant, A. H., 1942, Класифікація роду Drosophila, з описами дев'яти нових видів. Univ. Texas Publ., 4213: 6-51.
- Tidon-Sklorz, R. & Sene, F. M., 1992, Вертикальний та тимчасовий розподіл видів Drosophila (Diptera, Drosophilidae) в лісовій місцевості штату Сан-Паулу, Бразилія. Rev. Bras. Biol., 52: 311-317.
- Tidon-Sklorz, R. & Sene, F. M., 1995, Фауна Drosophila (Diptera, Drosophilidae) в північній частині “Cadeia do Espinhaço”, штатів Мінас-Жерайс та Баїя, Бразилія: біогеографічні та екологічні аспекти. Iheringia, Sér. Zool., 78: 85-94.
- Tidon-Sklorz, R. & Sene, F.M., 1999, Drosophila. In: C. R. F. Brandão & E. M. Cancello (eds), Наземні безхребетні. Т. V. Біорізноманіття штату Сан-Паулу. Синтез знань на кінець XX століття, FAPESP, Сан-Паулу.
- Tidon-Sklorz, R. & Sene, F. M., 2001, Два нові види з набору родичів Drosophila serido (Diptera, Drosophilidae). Iheringia, Sér. Zool., 90: 141-146.
- Val, F. C. & Kaneshiro, K. Y., 1988, Drosophilidae (Diptera) з Estação Biológica de Boracéia, на узбережжі штату Сан-Паулу, Бразилія: Географічний розподіл. In: P. E. Vanzolini & W. R. Heyer (eds.), Матеріали семінару з неотропічних патернів розподілу, Академія бразильських наук, Ріо-де-Жанейро.
- Val, F. C. & Sene, F. M., 1980, Нововведений вид Drosophila в Бразилії. Pap. Avul. Dep. Zool. Sec. Agric., 33: 293-298.
- Val, F. C., Vilela, C. R. & Marques, M. D., 1981, Drosophilidae неотропічного регіону. In: M. Ashburner, H. L. Carson & J. N. Thompson (eds.), Генетика та біологія Drosophila, Академічний прес, Лондон.
- Vanzolini, P. E., 1981, Квазіісторичний підхід до природної історії диференціації рептилій у тропічних географічних ізолятах. Pap. Avul. Dep. Zool. Sec. Agric., 34: 189-204
- Vermeij, G. J., 1996, Порядок денний для інвазійної біології. Biol. Conserv., 78: 3-9.
- Vilela, C. R., 1983, Ревізія групи видів Drosophila repleta (Diptera, Drosophilidae). Rev. Bras. Entomol., 27: 114.
- Vilela, C. R., 1999, Чи колонізує Zaprionus indianus Gupta, 1970, (Diptera, Drosophilidae) наразі неотропічний регіон? D. I. S., 82: 37-39.
- Vilela, C. R., Pereira, M. A. Q. R. & Sene, F. M., 1983, Попередні дані про географічний розподіл видів Drosophila в межах морфокліматичних доменів Бразилії.II. Група repleta. Ciênc. Cult., 35: 66-70.
- Zar, J. H., 1999, Множинні порівняння. У: Біостатистичний аналіз. 4-е вид., Prentice Hall Inc., Нью-Джерсі.