Морфологія едеагуса як дискримінантний маркер у двох тісно пов'язаних кактососних видах Drosophila (Diptera; Drosophilidae) в Південній Америці
Машинний переклад
Оригінальна стаття написана мовою EN (посилання для прочитання) .
Анотація
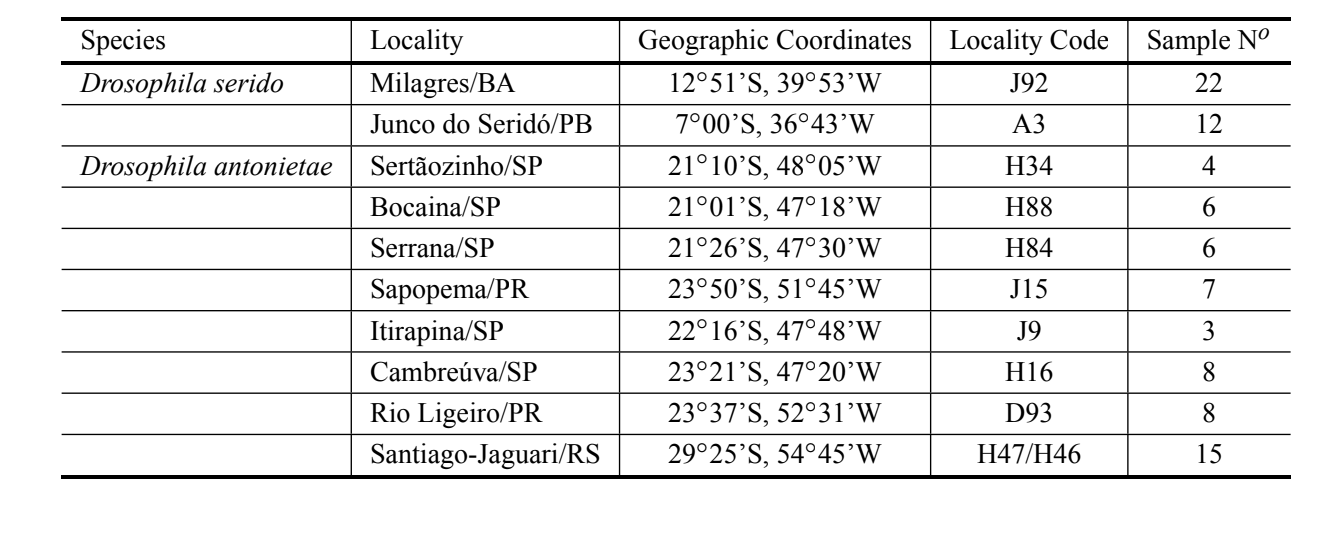
Drosophila serido та D. antonietae є спорідненими видами, що належать до кластеру Drosophila buzzatii . Морфологічно їх можна розрізнити лише за кількісними ознаками. У цій статті ми аналізуємо довжину та вирівняну середню кривизну чотирьох ділянок адеагуса D. antonietae та D. serido. Зразки D. serido та D. antonietae були правильно класифіковані 96.74% часу. Виходячи лише з ознаки, яка найбільше сприяла розрізненню груп (вирівняна середня кривизна арки IV адеагуса), ми спостерігали значну інтраспецифічну морфологічну дивергенцію у D. serido відносно D. antonietae, що узгоджується з іншими маркерами. Висока морфологічна дивергенція у вирівняній середній кривизні арки IV адеагуса показує, що ця ділянка еволюціонувала швидше, ніж інші, з моменту дивергенції двох видів. Обговорюється важливість даного дослідження для розуміння генетичної основи, яка контролює формування адеагуса у видів кластеру Drosophila buzzatii .
Вступ
Морфологічні характеристики історично використовувалися в еволюційних та таксономічних дослідженнях. Проте відсутність якісних морфологічних відмінностей серед споріднених видів ускладнює встановлення діагностичних морфологічних характеристик серед них. У багатьох випадках кількісні варіації морфометричних ознак є достатніми для розрізнення споріднених видів (Moreteau et al. 2003, Moraes et al. 2004).
Морфологія аедеагу (зовнішні чоловічі статеві органи) має надзвичайне значення в таксономічній характеристиці видів Drosophila (Vilela 1983, Silva and Sene 1991, Liu et al. 1996, Moreteau et al. 2003, Kullikov et al. 2004), а також в інших двокрилих. Кількісні варіації морфологічних характеристик аедеагу також використовувалися для розрізнення споріднених видів Drosophila (Silva and Sene 1991, Kullikov et al. 2004). У групі Drosophila repleta , яка включає більше 90 неотропічних видів, більшість вважаються спорідненими видами. Проте існує помітна різниця у формі аедеагів (форма + розмір) серед них, що вважається основною діагностичною ознакою в групі (Vilela 1983).
Кластер Drosophila buzzatii (repleta група, mulleri підгрупа, buzzatii комплекс) є монофілетичною групою, що складається з семи споріднених кактусофільних видів: D. buzzatii, D. borborema, D. koepferae, D. antonietae, D. gouveai, D. serido та D. seriema. У цьому кластері кількісні відмінності морфологічних характеристик крила (Moraes et al. 2004) та чоловічих статевих органів (Silva and Sene 1991, Tidon-Sklorz and Sene 1995, Prado et al. 2004) успішно використовувалися для розрізнення видів і популяцій. Серед семи видів цього кластера D. buzzatii та D. borborema мають адеаги, які відрізняються за формою та розміром від інших видів (Tidon-Sklorz and Sene 1995). Однак морфологія адеагів інших видів кластера подібна до так званого типу адеагу D. serido (Рисунок 1a), і на основі цього спостереження ці види спочатку були класифіковані як такі, що належать до одного виду (Vilela and Sene 1977). Для вивчення адеагу цих видів Silva і Sene (1991) поділили його на чотири основні області, обмежені орієнтирами (Рисунок 1b). Кожна область, обмежена двома сусідніми орієнтирами, була названа аркою. На основі довжини арок дослідники визначили п'ять різних типів адеагу, названих від A до E, специфічних для кожного виду.
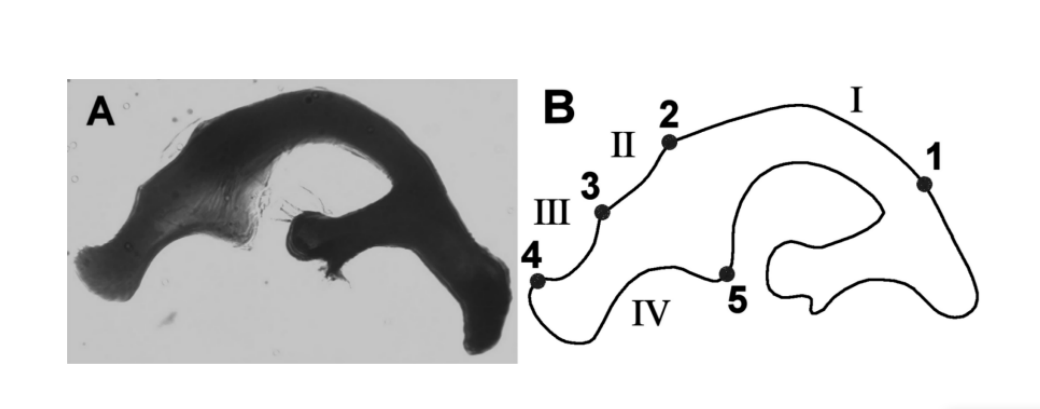
Drosophila serido та D. antonietae є двома спорідненими видами, що належать до кластеру Drosophila buzzatii , проаналізованого в цьому дослідженні. D. antonietae зростає в мезофільних лісах регіонів басейну Парана-Парагвай у Південній Америці, асоційованих з кактусом Cereus hildemannianus (Tidon-Sklorz і Sene 2001). D. serido присутній на північному сході Катаїнги та Атлантичному узбережжі Бразилії, від північного сходу до південного регіону Бразилії, асоційованих з рядом родів кактусів (Pereira et al. 1983). На південному межі ареалу D. serido існує зона контакту з D. antonietae (A.C. Morales et al., неопубліковані дані). Незважаючи на те, що D. serido та D. antonietae мають різні метафазні пластини (Baimai et al. 1983), алоензимні патерни (A.C. Morales et al., неопубліковані дані, Mateus і Sene 2003), мітохондріальний гаплотип (Manfrin et al. 2001) та супутнє ДНК (Kuhn і Sene 2005), вони мають одну й ту ж фіксовану хромосомну інверсію 2x7 (Ruiz et al. 2000), що може вказувати на тісний філогенетичний зв'язок. У морфологічному плані лише кількісні варіації можуть розрізняти ці два види (Silva і Sene 1991, Moraes et al. 2004).
У цій статті були проаналізовані морфометричні характеристики адеагуса зразків D. antonietae з кількох різних популяцій, що охоплюють усю відому територію поширення виду, а також зразків D. serido, що походять з двох різних популяцій. Ми обрали ці види, оскільки їх можна диференціювати за генетичними маркерами і, маючи вторинну контактну зону, з популяцією в симпатрії, з можливим утворенням гібридів у південному регіоні Бразилії. Окрім вимірювань довжини арки адеагуса, вже використаних у Silva і Sene (1991), також були використані вимірювання кривизни цих арок (Prado et al. 2004). Нашими основними цілями було ввести та протестувати використання вимірів кривизни у видів кластеру Drosophila buzzatii та визначити області адеагуса, які найбільше сприяли дискримінації видів D. serido та D. antonietae. Обговорювані тут дані підкреслюють важливість цих областей адеагуса для майбутніх досліджень з метою зрозуміти генетичну основу адеагуса видів, що належать до кластеру Drosophila buzzatii, а також для досліджень зони контакту між двома видами.
Матеріали та методи
Зразки
Було проаналізовано п'ятдесят вісім аедеаг з видів D. antonietae з кількох різних популяцій, що представляють всю їхню дистрибуцію, та 34 аедеаг D. serido, що належать до популяцій, розташованих у Junco do Seridó/PB та Milagres/BA (типова локалізація) (Таблиця I). Усі проаналізовані особини були зібрані в природних умовах (дикі мухи). Аедеаги були підготовлені у зрізах для оптичної мікроскопії відповідно до Канеширо (1969). Усі аедеаги, використані в цій статті, були депоновані в Лабораторії еволюційної генетики при USP – Рібейран-Прету. Зображення аедеаг були збільшені в 200 разів і оцифровані з мікроскопа (Axioplan2 Zeiss), оснащеного системою цифрового захоплення зображень Axiovision Zeiss та збережені на комп'ютері.
Вимірювання та статистичний аналіз
П'ять точок були визначені вручну як орієнтири, відповідно до Сілви та Сене (1991). Орієнтири були використані для встановлення внутрішніх контурів досліджуваної області; області між двома сусідніми орієнтирами є арками (Рисунок 1b).
Як у Сілви та Сене (1991), була виміряна довжина арок I, II, III та IV адеагуса (Таблиця II). Це вимірює відстань, накопичену між послідовними орієнтирами. Крім цього, ми також виконали вирівняні вимірювання кривизни арок. Коротко кажучи, безперервна кривизна є геометричною мірою, яка виражає швидкість зміни кута між дотичною до кривої та віссю x (Коста та ін. 2004). Значення вирівняної кривизни генеруються шляхом стандартизації за допомогою сигмоїди безперервної кривизни для підсилення точок з низькою кривизною в області інтересу (Коста та Сезар 2000). Цей етап дозволяє максимізувати інформацію про кривизну аналізованої структури. Для проведення аналізів ми використовували середні значення вирівняних кривизн арок I, II, III та IV адеагуса (Таблиця II). Вимірювання були отримані відповідно до методології, описаної Прадо та ін. (2004).
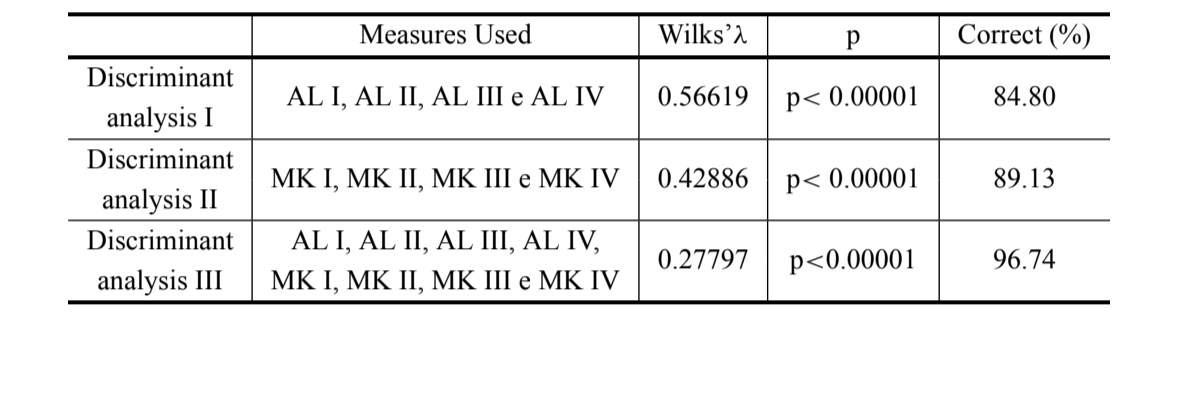
Ми виконали три різні дискримінантні аналізи, використовуючи лише виміри довжини арки (дискримінант I) та вирівняну середню кривизну (дискримінант II), а також дискримінантний аналіз, використовуючи всі виміри разом (дискримінант III). Ці аналізи були виконані для отримання правильних відсотків класифікації індивідів, щоб перевірити, які змінні більше сприяли дискримінації груп, і щоб порівняти використання вимірів кривизни з вимірами довжини арки, раніше виконаними Сілвою та Сене (1991). Нижчі значення лямбда Вілкса (які варіюють від 0 до 1) вказують на кращий аналіз. Крім цього, з даними дискримінантного аналізу III було виконано аналіз канонічних змінних. Аналіз дискримінантних та канонічних змінних був виконаний з використанням програмного забезпечення Splus (версія 1.8).
Абсолютне значення канонічних стандартизованих коефіцієнтів, отриманих внаслідок аналізу канонічних змінних, представляє вагу кожної змінної у формуванні канонічного кореня, де вищі значення відповідають найбільшій диференціації між групами. Ці коефіцієнти будуть представлені для показу відносного внеску кожної змінної у формування отриманого канонічного кореня та для визначення, яка змінна є найважливішою для дискримінації груп.
Окрім багатовимірного аналізу, найважливіша міра для дискримінації видів використовувалася індивідуально для отримання правильного відсотка класифікації індивідів за допомогою байєсівського методу класифікації (Duda et al. 2000). Для цього були встановлені нормальні криві на основі середнього значення та стандартного відхилення кожної групи стосовно змінної, визначаючи функцію щільності та ймовірності, що використовуються в байєсівському аналізі.
Результати
Результати трьох проведених дискримінантних аналізів наведені в таблиці III. Дискримінація між D. antonietae та D. serido є вищою, коли використовуються довжини арки адеагуса разом з вимірами кривизни (дискримінант III) (λ Вілкса = 0.27797; p<0.00001), де 96.74% індивідів класифікуються правильно (таблиця III), правильна класифікація D. antonietae становить 98.30%, а D. serido - 94.10%.
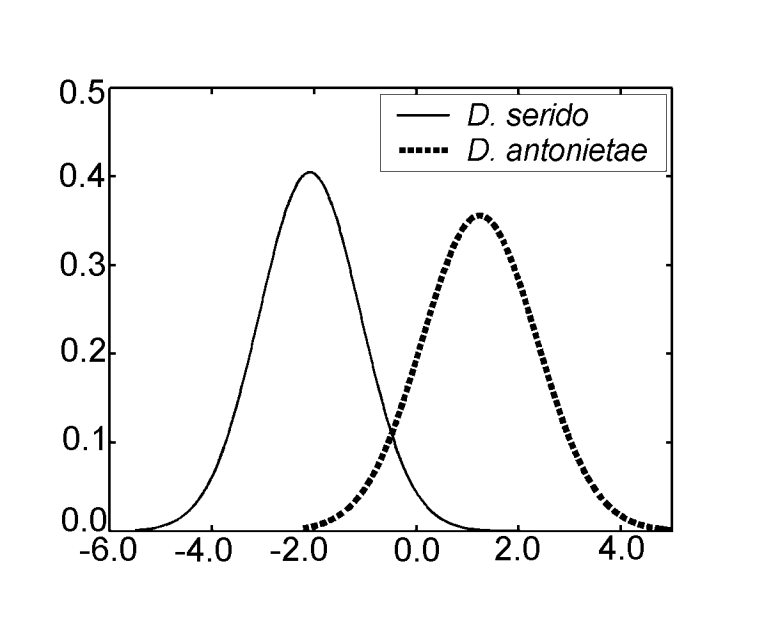
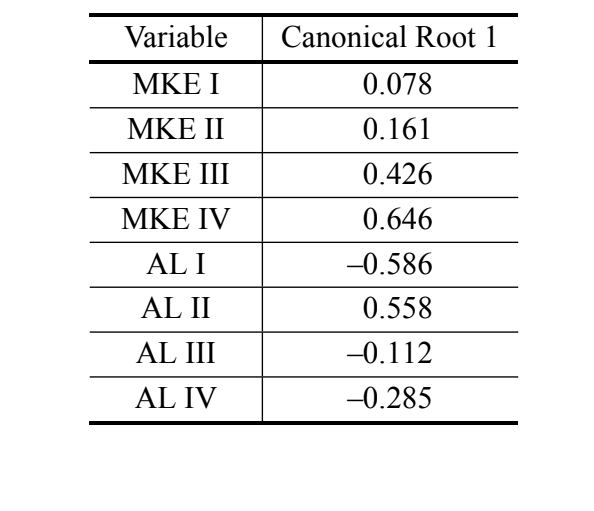
В аналізі канонічних змінних за допомогою заходів дискримінантного аналізу III було отримано лише одну канонічну вісь. Канонічні стандартизовані коефіцієнти (вага кожної змінної) наведені в Таблиці IV. Абсолютні значення цих коефіцієнтів вказують на те, що найважливішою змінною для формування канонічного кореня була MKE IV, тоді як найменш важливою була змінна MKE I. Нормальні криві, отримані за середнім значенням та стандартним відхиленням двох груп стосовно канонічних оцінок, показані на Рисунку 2, на якому ми можемо спостерігати дискримінацію між D. antonietae (вищі канонічні оцінки) та D. serido (нижчі канонічні оцінки).
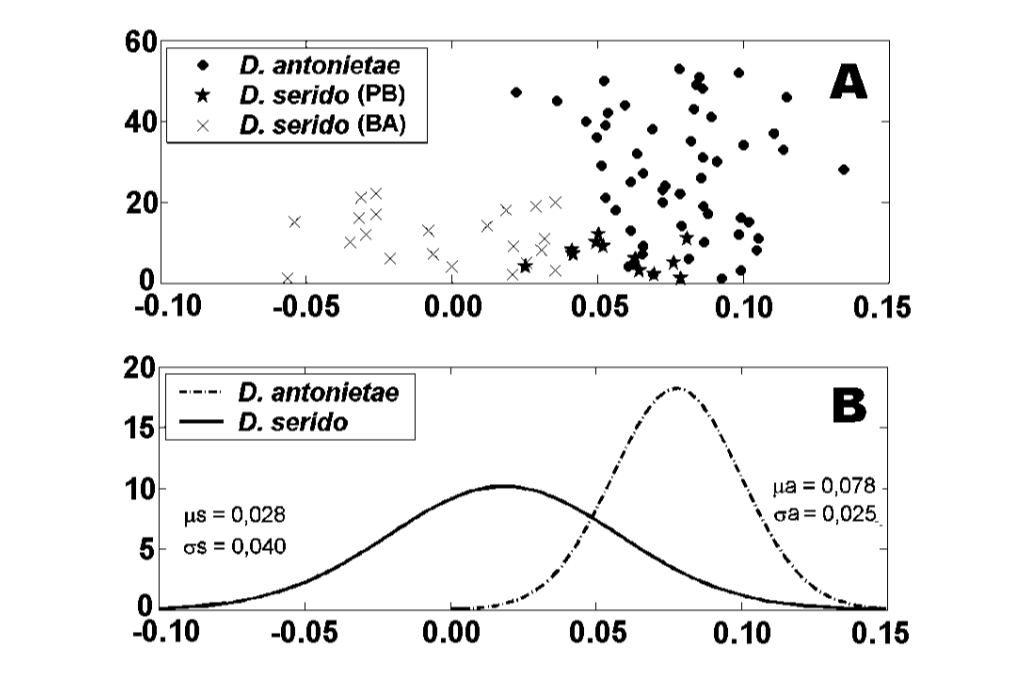
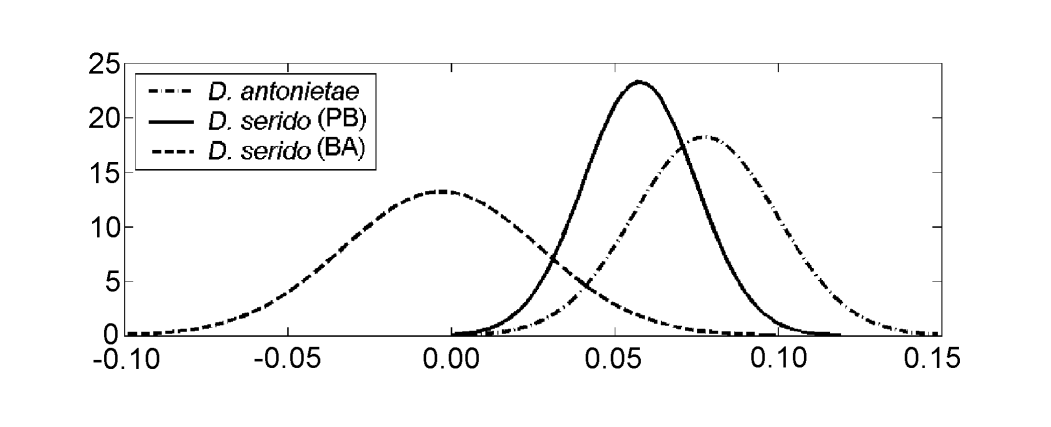
Значення для кожної особини, отримані з аналізу MKE IV, показані на малюнку 3a. Щодо цього показника, D. serido має вищу варіацію навколо середнього (середнє квадратичне відхилення = 0.04), ніж D. antonietae (середнє квадратичне відхилення = 0.025) (малюнок 3b), головним чином через те, що існують значні відмінності щодо цього показника між обома популяціями, які складають вибірку D. serido (малюнок 3a; малюнок 4).
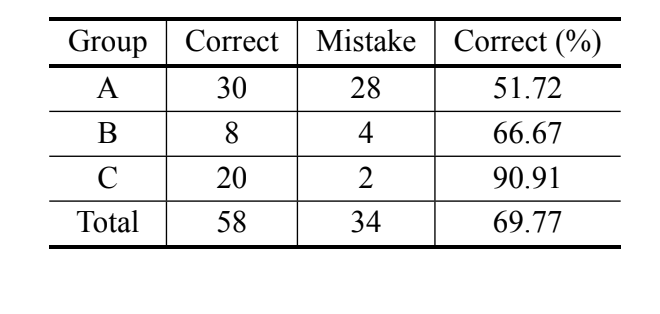
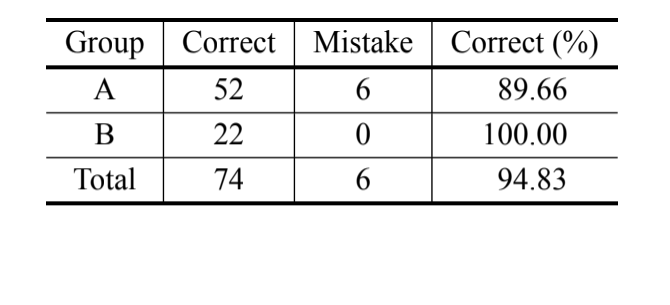
Відповідно до байєсівського аналізу, базуючись лише на вимірі MKE IV, 69.8% осіб були правильно класифіковані, і понад 90% осіб D. serido з Milagres/BA були правильно класифіковані. Проте, було зафіксовано високий відсоток помилок серед осіб D. serido з Junco do Seridó/PB та
D. antonietae (Таблиця V). Той же аналіз був проведений без осіб D. serido з Junco do Seridó/PB, і відсоток правильних класифікацій становив 94.83% (Таблиця VI), що показує екстремальну розбіжність між особами популяції D. serido з Milagres/BA та D. antonietae на основі MKE IV.
Обговорення
Наші дані підтверджують існування кількісних відмінностей у морфологічних характеристиках адеагуса проаналізованих особин, що дозволяє відрізнити D. serido від D. antonietae. Крім того, вирівняна середня кривизна арки IV (MKE IV) була змінною, яка найбільше сприяла дискримінації груп (Таблиця IV).
Довжина перших чотирьох арок адеагуса вже успішно використовувалася для розрізнення видів кластеру Drosophila buzzatii, серед яких D. serido та D. antonietae (адеагус типу A та D відповідно у Сілва і Сене 1991). У цій статті ми виявили значне підвищення розрізнення груп, додавши вимірювання вирівняної середньої кривини в морфометричному аналізі, де 96.74% індивідів правильно класифіковані (Таблиця III). Таким чином, вимірювання вирівняної середньої кривини є важливими для розрізнення D. antonietae та D. serido і можуть бути корисними в дослідженні зони симпатрії між цими видами в південному регіоні Бразилії. Оскільки вимірювання кривини не враховують розмір структури, можна припустити, що невеликі кількісні зміни в формі адеагуса, особливо в регіоні арки IV, могли відбутися з моменту кладогенезу видів D. serido та D. antonietae.
Існує значна морфологічна дивергенція стосовно MKE IV між D. antonietae та D. serido, оскільки D. antonietae показує відносно меншу варіацію навколо середнього, ніж D. serido (Рисунок 3b). Було проаналізовано особини з різних популяцій, що представляють всю територію розподілу популяцій D. antonietae (Таблиця I); у цьому зразку дані свідчать про те, що існує однорідність серед популяцій D. antonietae . Монтейро та Сене (1995) проаналізували морфологію аедеагуса особин з кількох популяцій D. antonietae за допомогою морфометричного методу тросової мережі і також не спостерігали морфологічних відмінностей серед них. Аллоензиматичні (Матейуш і Сене 2003) та мікросателітні патерни (Л.П.Б. Мачадо та ін., неопубліковані дані) також показали однорідність серед різних популяцій D. antonietae. Ці результати свідчать про генетичний потік між популяціями D. antonietae, що сприяється асоціації цього виду з його господарським кактусом Cereus hildmaniannus , який зустрічається в мезофільних галерейних лісах вздовж річок басейну Парани-Парагваю, що формують коридори міграції для особин виду D. antonietae (Монтейро та Сене 1995).
Drosophila serido має, в порівнянні, вищу міжвидову варіацію стосовно MKE IV, ніж D. antonietae (Рисунок 3b), як можна спостерігати в різницях, виявлених серед особин з популяцій Junco do Seridó/PB та Milagres/BA. Особини D. serido з популяції Junco do Seridó/PB мають проміжні значення для MKE IV між тими, що виявлені для популяцій D. serido в Milagres/BA та популяцій D. antonietae, які були проаналізовані (Рисунок 4). Ці дані свідчать про морфологічну диференціацію між обома популяціями D. serido, ймовірно, через обмеження генетичного потоку між ними. Між бразильськими штатами Баїя та Пернамбуко знаходиться велика річка, річка Сан-Франсиску, яка може діяти як географічний бар'єр, що перешкоджає генетичному потоку між популяціями D. serido, проаналізованими в цій статті. Морфологічні розбіжності серед популяцій D. serido узгоджуються з іншими роботами, оскільки цей вид є політиповим стосовно інших маркерів. Північно-східні популяції D. serido мають метафазні пластини типу I, тоді як деякі прибережні популяції мають метафазні пластини типу III (Arraial do Cabo/RJ) та типу IV (Peruíbe/SP) (Baimai et al. 1983). Що стосується хромосомних інверсій, хоча всі популяції D. serido мають фіксовану інверсію 2x7, чотири поліформні інверсії обмежені північно-східними популяціями (2a8, 2b8, 2c8 та 2d8) і дві фіксовані інверсії (2x8 та 2w8) зустрічаються в прибережних популяціях (Tosi and Sene 1989, Ruiz et al. 2000). Крім того, популяції D. serido з північно-східного Бразилії частково ізольовані репродуктивно від популяції Arraial do Cabo/RJ вздовж бразильського узбережжя (N.M.V. Bizzo, неопубліковані дані). Недавні дослідження також свідчать про дискримінацію серед популяцій D. serido на північному сході та прибережному Бразилії на основі диференціації мітохондріальних гаплотипів (A.C. Morales et al., неопубліковані дані).
Різноманітність серед популяцій одного й того ж виду є важливою вимогою в процесі видоутворення, тому дослідження, що охоплюють різноманітні популяції політипового виду, такого як D. serido, є важливими для кількісної оцінки морфологічної дивергенції у зв'язку з генетичною дивергенцією в природних популяціях.
Не всі арки адеагуса однаково інформативні для розрізнення видів. Згідно з дослідженнями Сілви та Сене (1991), арки II та III (Рисунок 1) є найбільш інформативними для розрізнення видів у кластері Drosophila buzzatii. У порівнянні парами, використовуючи метод тросової мережі, вимірювання, пов'язані з аркою III, були найзначнішими для розрізнення між
D. antonietae та D. gouveai (Монтейро та Сене 1995). Через аналіз геометричної морфометрії, використовуючи вимірювання кривизни, Прадо та ін. (2004) спостерігали, що кривизна та нормалізована довжина арки III є важливими вимірюваннями для розрізнення між видами D. gouveai та D. antonietae. Наші дані показують, що MKE IV є найважливішим для розрізнення видів D. serido та D. antonietae; і, спираючись лише на це вимірювання, можливо розрізнити більше 94% індивідів D. serido з популяції Мілагрес/BA (типова локалізація) від тих, що належать до D. antonietae (Таблиця VI). Ці результати показують, що дистальна частина адеагуса (арка III + арка IV) має найбільші міжвидові відмінності, незалежно від інших частин цього органу, що свідчить про те, що цю область можна вважати "гарячою еволюційною точкою" (Куліков та ін. 2004) для адеагуса під час подій кладогенезу в кластері Drosophila buzzatii, але це питання все ще залишається відкритим для обговорення.
Морфометричні варіації між групами організмів та всередині них відображають вираження фенотипу, що є результатом інтегрованого полігенного контролю, який змінюється під час кладогенезу та еволюції груп (Falconer 1989). Крім того, кілька епігенетичних та екологічних факторів можуть впливати на формування структури (Atchley et al. 1992), що ускладнює ідентифікацію причин морфологічної дивергенції серед популяцій та видів. Проте, порівняльні морфометричні дослідження гомологічних регіонів обох видів, таких як арки адеагуса, є важливими вимогами для розуміння генетичної основи, яка контролює формування цього регіону. Хоча генетична основа адеагуса виявляється для інших груп Drosophila (Liu et al. 1996), гени, які контролюють формування адеагуса у видів кластеру Drosophila buzzatii , ще не встановлені. Високий відсоток правильності індивідів, проаналізованих у цій статті, що базується виключно на MKE IV, вказує на те, що регіон арки чотири може бути корисним для майбутніх досліджень з розуміння генетичної основи адеагуса у цих видах, оскільки встановлення інформативних морфологічних характеристик серед різних таксономічних груп є попереднім кроком у вивченні QTL (Кількісні локуси ознак), методології для виявлення, картування та оцінки впливу деяких локусів на метричні характеристики.
Автори: Фернандо Ф. Франко, Педро Р.Р. Прадо, Фабіо М. Сене, Лучіано Ф. Коста та Маура Х. Манфрін
Посилання
- Atchley WR, Cowley DE, Vogi C та Mc-Lellan T. 1992. Еволюційна дивергенція, зміна форми та генетична кореляційна структура в щелепі гризунів. Syst Biol 41: 196–221.
- Baimai V, Sene FM та Pereira MAQR. 1983. Гетерохроматин та каріотипічна диференціація деяких неотропічних видів кактусів з групи Drosophila repleta. Genetica 67: 81–92.
- Costa LF та Cesar RMJ. 2000. Аналіз форми та класифікація: теорія та практика. CRC Press Book Series on Image Processing. Cambridge Univ Press, New York, 659 с.
- Costa LF, dos Reis SF, Arantes RAT, Alves ACR та Multinari G. 2004. Біологічний аналіз форми за допомогою цифрової кривизни. Pattern Recognit 37: 515–524.
- Duda RO, Hart PE та Stork DG. 2000. Класифікація шаблонів. J Wiley & Sons, New York, USA, 680 с.
- Falconer DS. 1989. Вступ до кількісної генетики. 2-е вид., Лондон: Longman.
- Kaneshiro KY. 1969. Дослідження взаємовідносин видів Hawaiian Drosophila на основі зовнішніх чоловічих статевих органів. Univ Texas Publ 6918: 55–70.
- Kuhn GCS та Sene FM. 2005. Еволюційний обіг двох підродин супутникової ДНК pBuM у кластері видів Drosophila buzzatii (група repleta): від alpha до alpha/beta масивів. Gene 349: 77–85.
- Kullikov AM, Melnikov AI, Gornostaev NG, Lazebny OE та Mitrofanov VG. 2004. Морфологічний аналіз статевого органу самців у групі видів Drosophila virilis: багатовимірний підхід. J Zool. Syst Evol Res 42: 135–144.
- Liu J, Mercer JM, Stam LF, Gibson GC, Zeng Zhao-Bang та Laurie CC. 1996. Генетичний аналіз морфологічної різниці у статевих органах самців Drosophila simulans та D. mauritiana. Genetics 142: 1129–1145.
- Manfrin MH, Brito ROA та Sene FM. 2001. Систематика та еволюція кластеру Drosophila buzzatii (Diptera; Drosophilidade) за допомогою mtDNA. Ann Entomol Soc Am 94: 333–346.
- Mateus RP та Sene FM. 2003. Тимчасова та просторово-алозимна варіація у південноамериканському кактусоносному Drosophila antonietae (Diptera; Drosophilidae). Biochem Genet 41: 219–233.
- Monteiro SG та Sene FM. 1995. Морфометричне дослідження популяцій Drosophila serido з Центрального та Південного Бразилії. Rev Bras Genet 18 (Suppl.): 283.
- Moraes EM, Spressola VL, Prado PRR, Costa LF та Sene FM. 2004. Дивергенція в морфології крил серед споріднених видів кластеру buzzatii. J Zool Syst Evol Res 42: 154–158.
- Moreteau B, Gibert P, Pétavy G, Moereteau JC, Huey RB та David JR. 2003. Морфометрична еволюція в кладі Drosophila: група Drosophila obscure. J Zool Syst Evol Res 41: 64–71.
- Pereira MAQR, Vilela CR та Sene FM. 1983. Нотатки про місця розмноження та живлення деяких видів групи repleta роду Drosophila (Diptera; Drosophilidae). Cienc Cult 35: 1313–1319.
- Prado PRP, Franco FF, Manfrin MH, Costa LF та Sene FM. 2004. Легкий та швидкий спосіб аналізу морфометричних характеристик. Proc Third Braz Symp Mathem Comp Biol 1: 329–340.
- Ruiz A, Cassian AM, Kuhn GCS, Alves Maqr та Sene FM. 2000. Головоломка сп Speciation Drosophila serido: складання нових частин разом. Genetica 108: 217–227.
- Silva AFG та Sene FM. 1991. Морфологічна географічна варіабельність у Drosophila serido (Diptera; Drosophilidae). Rev Bras Entomol 35: 455–468.
- Tidon-Sklorz R та Sene FM. 1995. Еволюція кластеру buzzatii (Drosophila група видів repleta) у середній Південній Америці. Evolución Biologica 9: 71–85.
- Tidon-Sklorz R та Sene FM. 2001. Два нові види з набору споріднених Drosophila serido (Diptera; Drosophilidae). Iheringia 90: 141–146.
- Tosi D та Sene FM. 1989. Подальші дослідження хромосомної варіабельності в складному таксоні Drosophila serido (Diptera; Drosophilidae). Rev Bras Genet 12: 729–745.
- Vilela CR. 1983. Ревізія групи видів Drosophila repleta (Diptera; Drosophilidae). Rev Bras Entomol 27: 1–114.
- Vilela CR та Sene FM. 1977. Два нові неотропічні види групи repleta роду Drosophila (Diptera; Drosophilidae). Papéis Avulsos de Zoologia 30: 295–299.