Генетична варіація гавайських дрозофіл. IV. Алозимна схожість між D. silvestris та D. heteroneura з острова Гаваї
Машинний переклад
Оригінальна стаття написана мовою EN (посилання для прочитання) .
Анотація
Види є ендеміками найновішого острова архіпелагу і широко симпатричні. Вони легко відрізняються морфологічно у обох статей. Використовуючи стандартні електрофоретичні процедури, ми дослідили 25 локусів, що кодують структурні білки, з 539 silvestris та 325 heteroneura , зібраних у трьох широко розділених місцях, де два види є симпатричними. Паралельні порівняння всередині та між видами показують наступні коефіцієнти генетичної ідентичності (Nei's I): всередині silvestris 0.961 ± 0.01; всередині heteroneura 0.949 ± 0.02; між silvestris та heteroneura, 0.939 ± 0.01. Ні відмінності всередині, ні між видами не є значущими. Немає фіксованих алельних відмінностей ні всередині, ні між видами. У трьох зонах симпатії види показують різниці частот генів P < 0.05) на 9, 11 та 13 локусах відповідно. Це не дуже відрізняється від варіації всередині будь-якого з видів у трьох місцях. Обидва види мають подібні рівні гетерозиготності (H) (silvestris, 0.083; heteroneura, 0.089) та відсоток поліморфних локусів (обидва 0.37). Пропонується, що незважаючи на їх морфологічну дивергенцію, ці види є набагато новішими, ніж класичні братні види. Значні алозимні відмінності, можливо, не встигли накопичитися.
DROSOPHILA silvestris та D. heteroneura є парою тісно пов'язаних гігантських видів, які є ендеміками лише тропічних лісів острова Гаваї, найновішого в архіпелазі. Вони симпатричні в більшості районів, за винятком вищих висот, де популяції, як правило, є виключно silvestris.
Види легко відрізняються морфологічно (Hardy 1965) не лише завдяки надзвичайно широкій голові обох статей heteroneura, але й за кольоровими малюнками обличчя, грудей, плеври та черевця, а також за відмінностями в малюнку крил. D. silvestris має шість унікальних інверсій у поліморфному стані; сьома інверсія є поліморфною в обох видах (Carson and Stalker 1968; Craddock and Carson 1975). Між видами немає фіксованих інверсій, і обидва мають спільний набір стандартних послідовностей, тобто є хромосомно гомосеквенційними.
Хоча між видами існує сильна статева ізоляція в лабораторних схрещуваннях (Ahearn et al. 1974; Kaneshiro 1976), гібриди F1 були отримані з обох взаємних схрещувань (Craddok 1974; Ahearn і Val 1975). Обидві статі цих гібридів F1 є повністю плідними, і генетична сегрегація морфологічних відмінностей між видами була спостережена в поколінні F, та в зворотних схрещуваннях (Val 1976).
Попередні дослідження алозимів показали надзвичайну схожість між двома видами (S = 0.96; Johnson et al. 1975; Johnson і Carson 1975). У даній статті розширено дослідження генетичної варіабельності всередині та між двома видами на розчинні білки, закодовані 25 локусами. Дані отримані з трьох областей симпатрії. Результати підтверджують вражаючу біохімічну схожість видів, незважаючи на генетичні відмінності між ними в морфології, інверсіях та поведінці.
Матеріали та методи
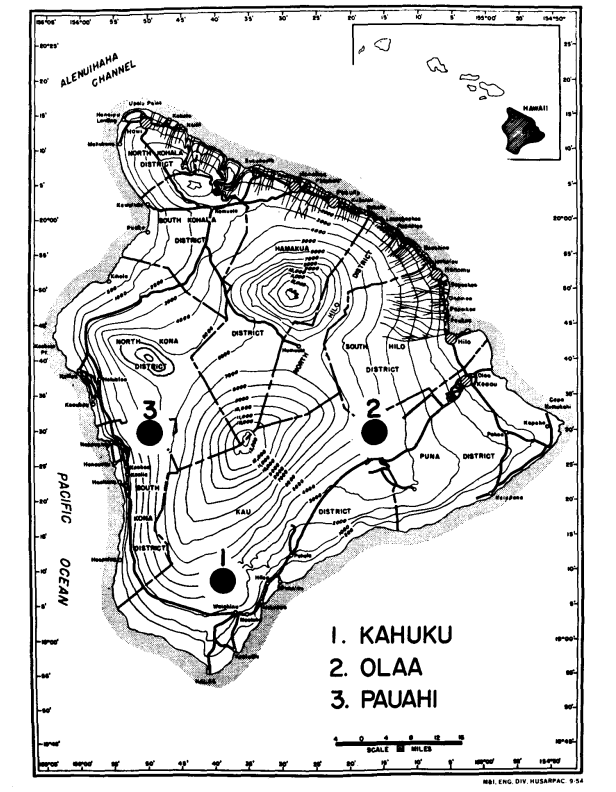
У 1974 та 1975 роках ми зібрали два види з трьох районів помірної висоти на Гаваях (Великий острів), де вони симпатричні. Це (Рисунок 1): (1) ранчо Кахуку в південній частині округу Кау, (висота 1158 м); (2) Олаа, ліс біля вулканічної експериментальної станції на дорозі Райт, округ Пуна (висота 1238 м) та (3) Пауахі, округ Південний Кона (висота 1311 м). Новоуловлені самці використовувалися для електрофорезу якомога швидше після улову. Самки були ізольовані та дозволено їм виробляти потомство F1 для використання в приготуваннях слинних залоз. Після виробництва личинок матері використовувалися для електрофорезу. Тільки черевце мух підлягало електрофорезу; решта (голова, грудна клітка та придатки) була змонтована та поміщена в колекцію для використання в морфологічних аналізах.
Після видалення живіт був заморожений на ніч, а потім індивідуально гомогенізований у 0,05 мл демінералізованої дистильованої води, і кожен супернатант абсорбувався чотирма сірниками фільтрувального паперу Whatman No. I (1 X 0,2 см). Ці сірники були потім розділені та застосовані до чотирьох горизонтальних крохмальних гелів, що поєднують різні буферні системи. Після електрофорезу кожен гель був нарізаний горизонтально чотири або п’ять разів, і ці скибочки фарбувалися окремо для одного або кількох з 25 ферментів. Уся робота виконувалася з використанням Electrostarch партії 303 при концентрації 12%.
Три комбінації буферів, використані в дослідженні, такі: Система буферів A = Гель-буфер: pH 8.9, 0.0076 м Трис, 0.005 м цитрат; Буфер електродів: pH 8.7, 0.269 м борат, 0.1 м натрію гідроксид. Система буферів C = Гель-буфер: pH 8.1, 0.074 м Трис, 0.008 м цитрат; Буфер електродів: pH 8.1, катод = 0.343 м Трис, 0.079 м цитрат; анод = 0.458 м Трис, 0.0104 м цитрат. Система буферів JRP = Гель-буфер: pH 7.0, 0.009 м Трис, 0.003 м цитрат; буфер електрода: 0.135 м Трис, 0.040 м цитрат. Усі ці буфери та методи фарбування, що використовуються, подібні до описаних Штайнером і Джонсоном (1973) та Аяла та ін. (1972; 1974a).
Алелі в певному локусі були пронумеровані відповідно до відносних мобільностей їх алозимів на зазначених електрофоретичних буферних системах. Алель, що виробляє найчастіший алозим, був позначений як l.00, а алелі, що відповідають швидшим або повільнішим мігруючим смугам, були довільно призначені значення, що вказують на їх відповідні мобільності.
Буфер, що використовується, та позначення для кожного ферменту є наступними: Буферна система A : Естераза (EST, 1 локус) ; Алкалінова фосфатаза (AP, 1 локус) ; Кисла фосфатаза (ACPH, 1 локус) ; Глутамат оксалоацетат трансаміназа (GOT, 2 локуси) ; Альдегід оксидаза (AO, 1 локус) ; Октанол дегідрогеназа (ODH, I локус) ; Алкогольна дегідрогеназа (ADH, 1 локус) ; Ксантин дегідрогеназа (XDH, 1 локус) ; Лейцин амінопептидаза (LAP, 2 локуси) ; Гліцеральдегід-3-фосфат дегідрогеназа (G-3-PDH, 1 локус). Буферна система C : Малатна ензима (ME, 1 локус) ; Малат дегідрогеназа (MDH, 2 локуси) ; α-Гліцерофосфат дегідрогеназа (α-GPDH, 1 локус) ; Фосфоглюкомутаза (PGM, I локус) ; Фумараза (FUM, 1 локус) ; Сорбітол дегідрогеназа (SDH, 1 локус) ; Гексокіназа (HK, 2 локуси) ; Ізоцитрат дегідрогеназа (IDH, 1 локус). Буферна система JRP : 6-Фосфоглюконатна дегідрогеназа (6-PGDH, 1 локус) ; Гідроксибутират дегідрогеназа (HBDH, 1 локус) ; Фосфоглюкозоізомераза (PGI, 1 локус).
Результати
Основні дані алозимів наведені в таблиці 1. Таблиця 2 показує результати використання даних з таблиці 1 для розрахунку коефіцієнтів генетичної схожості, I, та відстані, D (Nei 1972) у парному порядку всередині та між видами. У тій же таблиці (нижче діагоналі) показано кількість локусів, які демонструють відмінності в частоті генів (P ≤ 0.05) у парних порівняннях. Розрахунок був виконаний за допомогою χ2, використовуючи частоту більш поширеного алеля проти об'єднаної частоти інших алелів. Використовувалася корекція Йейтса.
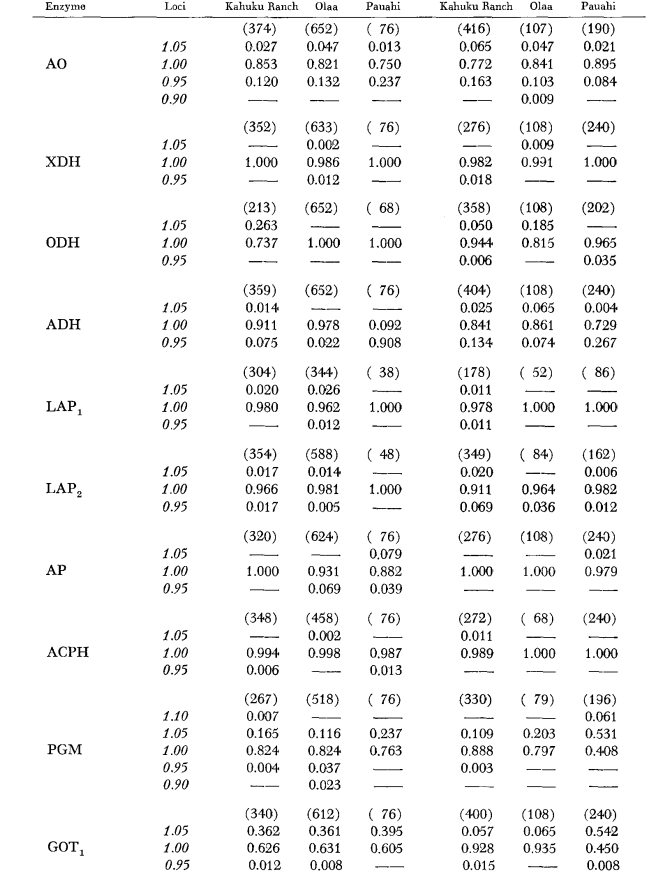
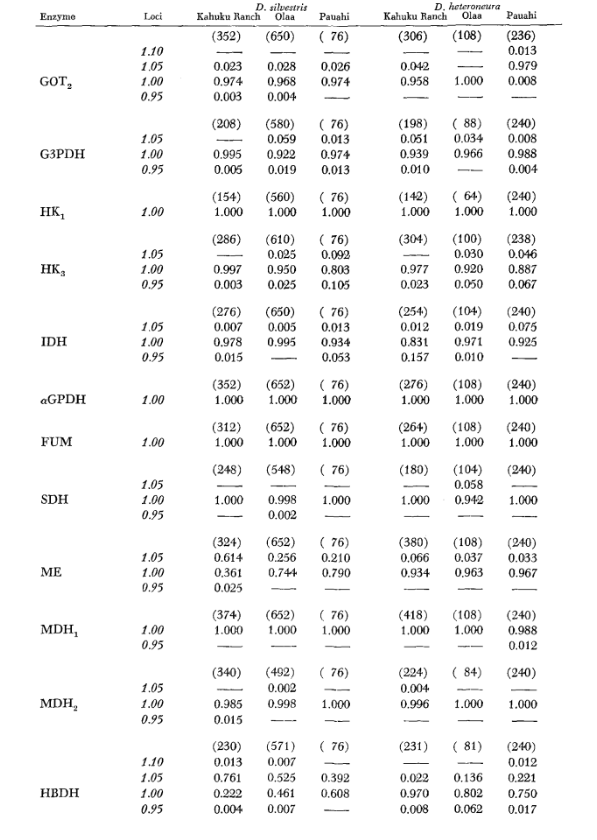
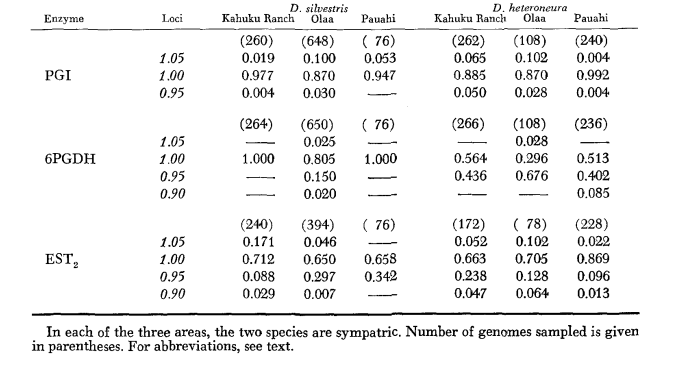
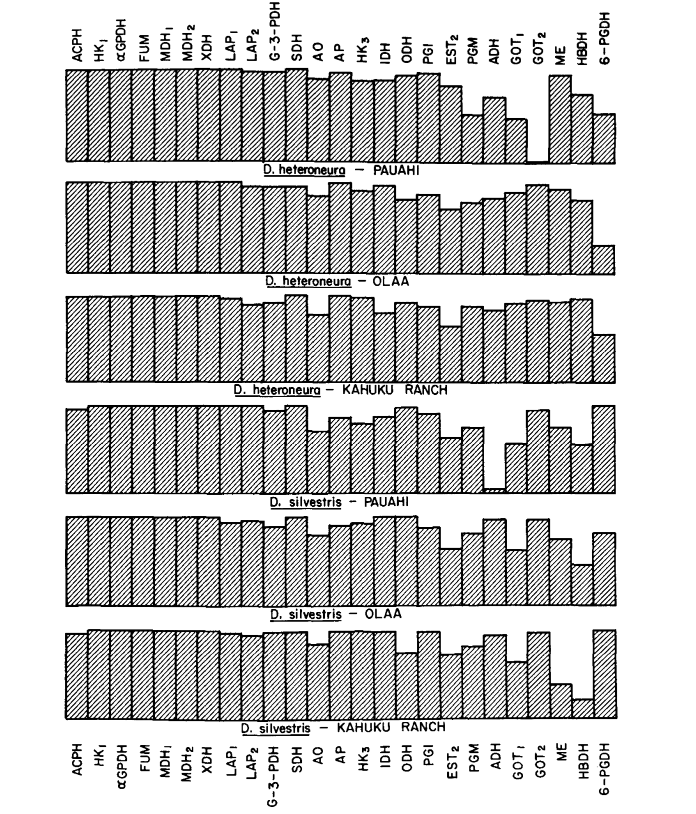
Середні рівні гетерозиготності та оцінки частоти поліморфних локусів наведені в Таблиці 3. Графічне представлення частоти алелю 1.0 для кожного алозиму в кожній популяції представлено на Рисунку 2.
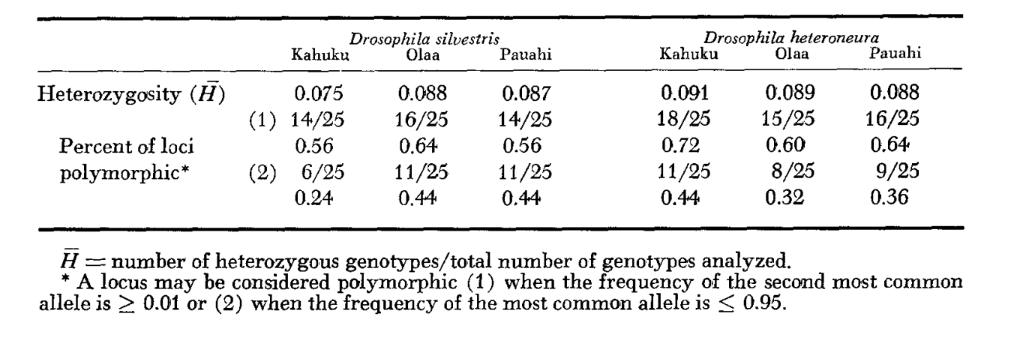
Алозимна схожість є дуже великою як всередині, так і між видами, незалежно від того, чи вимірюється вона коефіцієнтами схожості, чи різницями у частоті генів (див. резюме в Таблиці 4). Не існує фіксованих алельних відмінностей ні всередині, ні між видами. У трьох зонах симпатії види демонструють різниці у частоті генів (P = 0.05) на 9, 11 та 13 локусах відповідно. Це не дуже відрізняється від варіації всередині кожного з видів у трьох місцях (silvestris: 5, 7, 10; heteroneura: 7, 9, 12). Обидва види мають подібні рівні гетерозиготності (silvestris 0.083; heteroneura 0.089) та відсоток поліморфних локусів (обидва 0.37).

Деякі деталі є цікавими. Схожість між видами найменша в Pauahi (Таблиця 2; Рисунок 2). Перевірка показує, що це в основному пов'язано з відмінностями у частоті на ADH та GOT2. Великі відмінності на цих локусах спостерігаються між видами лише в цій області; тобто, 1.00 алель GOT2 низький лише в Pauahi heteroneura і 1.00 алель ADH низький лише в Pauahi silvestris (Таблиця 1; Рисунок 2).
Схожість між видами дещо більша в районі Кахуку. Тут відмінності стосуються переважно GOT1, ME та HBDH. У Олаа схожість між видами є найбільшою. GOT1 та HBDH знову показують відмінності, хоча вони не такі великі, як між видами в Кахуку.
Обговорення
Генетичні порівняння популяцій за допомогою алозимів заполонили літературу за останні десять років (див. огляди Айали 1975 року та Пауела 1975 року). Оскільки як досліджувані білки, так і електрофоретичні методи в цілому були подібними, результати з досить різних таксонів можуть бути корисно порівняні. Наприклад, генетична схожість між двома видами ящірок, коли зведена до коефіцієнта, такого як коефіцієнт Ней, може бути безпосередньо порівняна з подібними даними з пар видів мишей, рослин або Drosophila.
У своїй знаковій статті Левонтін і Хаббі (1966) вказали на ряд причин, чому стандартні електрофоретичні методи будуть схильні до заниження генетичної варіабельності та генетичної різниці. Це слід мати на увазі ще більше сьогодні через нещодавній розвиток послідовного електрофоретичного аналізу. Таким чином, Сінгх, Левонтін і Фелтон (1976) та Койн (1976) показали, що для локусу ксантиндегідрогенази D. pseudo-obscura та persimilis кількість алелів у досить малих природних зразках цих видів принаймні в чотири рази більша, ніж кількість, виявлена стандартними методами. Що стосується міжпопуляційних порівнянь, ці нові методи, здається, напевно зменшать спостережувані схожості на всіх рівнях диференціації популяцій (наприклад, місцеві популяції, підвиди, семісpecies, види).
Незважаючи на ці нові розробки, порівняння, засновані на стандартних методах, продовжують бути цінними, якщо їх обмеження усвідомлюються. D. silvestris та heteroneura є набагато ближчими алозимно, ніж будь-які інші пари видів Drosophila, які ще досліджувалися, включаючи всі пари "сibling" видів, підвидів та семісpecies (Ayala et al. 1974b). Насправді, коефіцієнти схожості для популяцій всередині або silvestris, або heteroneura мають той же порядок величини, що й ті, що між цими двома видами. Маленькі відмінності, які існують між парами популяцій двох видів, більше того, не є однаковими в трьох вивчених районах; тобто, вони не залучають однакові локуси або алелі.
Як обговорювалося у вступі, ми продовжуємо визнавати ці два утворення як хороші біологічні види. Вони морфологічно відмінні, широко симпатричні, поведінково ізольовані та різні у своїх хромосомних поліморфізмах. Морфологічні відмінності залежать від приблизно чотирнадцяти полігенів (Val 1976).
Вражаюча та важлива особливість цих видів, однак, полягає в тому, що реверсивні гібриди, отримані в лабораторії, є повністю фертильними в обох статях (Craddock 1974; Ahearn і Val 1975). Відповідно, природне гібридизацію слід враховувати. Хоча Craddock (1974) не знайшов жодних доказів цього, Kaneshiro і Val (1977) повідомили, що приблизно два відсотки диких мух, захоплених на ранчо Кахуку, можуть бути ідентифіковані як гібриди за морфологічними ознаками. Це той самий регіон, де були знайдені природні гібриди між двома іншими видами (Carson, Nair і Sene 1975). Морфометричний аналіз форми голови був успішно використаний для визначення точного реверсивного походження певних особин F, гібридів. В одному випадку дика самка D. silvestris виробила потомство, що складається виключно з F, гібридів. Оскільки кожна дика самка була ізольована від усіх самців відразу після захоплення, очевидно, що відповідні спарювання відбулися в природі. Слід підкреслити, що фон широких морфометричних досліджень лабораторних гібридів (Val 1976) підготував ґрунт для цих нових природних спостережень і, відповідно, діагностика цих випадків ґрунтується на надійних порівняльних доказах. Усі природні гібридні особини були виключені з електрофоретичних даних, представлених у цій статті.
Ще один факт, який додає до незвичайної ситуації щодо цих видів, - це їхня близька екологічна схожість. Обидва види були вирощені з одного й того ж індивідуального гниючого стебла загального рослинного господаря, Clermontia. Обидва, так само, формують свої лекси в дуже схожих місцях, так що самці та самки обох видів взаємодіють у природі.
Яка причина такої великої алозимної схожості цих видів? Дві можливості приходять на думку. По-перше, коли відбулося розходження між видами, це могло супроводжуватися певною кількістю електрофоретичного розходження. Таким чином, спостережувані в даний час схожості вимагали б, щоб такі відмінності були знищені інтрогресивною гібридизацією після утворення видів. Друга точка зору стверджує, що лише незначні електрофоретичні зміни супроводжували події видоутворення спочатку. Це може бути пов'язано з дуже значною екологічною близькістю.
Ми схиляємося до другої точки зору з ряду причин. Обидва види є найбільш схожими в Олаа. Проте, сильні докази нещодавньої гібридизації були отримані лише для Кахуку, де види є найбільш віддаленими алозимно. У Кахуку, крім того, природні популяції не дають жодних свідчень про формування "гібридного рою", що нагадує широке генетичне recombination, спостережуване в лабораторних схрещуваннях. Насправді, інтрогресія між видами, ймовірно, відбувається, але вона може бути обмежена вузьким генотипним спектром через природний відбір. Таким чином, лише гени біологічно незначного значення ( "відкрита система", Карсон 1975) можуть перетікати через бар'єр видів. Навпаки, інші гени можуть позначати або бути функціональною частиною закритої системи епістатичних балансів, унікальних для одного або іншого з видів. Таким чином, відбір може служити для запобігання розриву такої "закритої системи" (Карсон 1975).
Через геологічну новизну острова Гаваї, ми схильні припустити, що D, silvestris та D. heteroneura можуть справді бути новоствореними в часі. Ця обставина може бути основною причиною електрофоретичної схожості двох видів. Справді, вони можуть бути значно новішими, ніж класичні пари "сibling" видів (див. Carson 1976b). Як і в інших таких очевидних неосpecies, (наприклад, Gottlieb 1974; Turner 1974; Avise, Smith і Ayala 1975), алозимна диференціація була відносно незначною. Справді, існує можливість, що алозимна різниця може просто корелювати з часом, що минув з моменту кладастичного події, яка розділила два порівнювані об'єкти (Avise, Smith і Ayala 1975; Carson 1976a). Морфологічні відмінності, з іншого боку, можуть слугувати кращими індикаторами основних ефектів регуляторних генів, що характеризують важливі генетичні відмінності між новоствореними видами.
Автори: Ф. М. Сене, Х. Л. Карсон
Цитована література:
- Ахерн, Дж. Н., Х. Л. Карсон, Т. Добжанський та К. Й. Канешіро, 1974 Етологічна ізоляція між трьома видами підгрупи планітібія гавайської Drosophila. Proc. Natl. Acad. Sci. U.S. 71: 901—903.
- Ахерн, Дж. Н. та Ф. С. Вал, 1975 Плідні міжвидові гібриди двох симпатричних гавайських Drosophila. Генетика 80: s9.
- Авізе, Дж. С., Дж. Дж. Сміт та Ф. Дж. Аяла, 1975 Адаптивна диференціація з незначними генетичними змінами між двома рідними каліфорнійськими мінновими. Еволюція '29: 411—426.
- АЯЛА, Ф. Дж., 1975 Генетична диференціація під час процесу видоутворення. Еволюційна біологія 8: 1—78.
- Аяла, Ф. Дж., Дж. Р. Пауел, М. Л. Трейсі, К. А. Мурао та С. Перес-Салас, 1972 Варіабельність ферментів у групі Drosophila willistoni. IY Генетична варіація в природних популяціях Drosophila willistoni. Генетика 70: 113—139.
- Аяла, Ф. Дж., М. Л. Трейсі, Л. Г. Барр та Дж. Г. Еренфельд, 1974а Генетична та репродуктивна диференціація підвиду Drosophila equinoxialis caribbensis. Еволюція 28: 24—41.
- Аяла Ф. Дж., М. Л. Трейсі, Д. Хеджкок та Р. С. Річмонд, 1974б Генетична диференціація під час процесу видоутворення в Drosophila. Еволюція 28: 576—592.
- Карсон, Х. Л. 1975 Генетика видоутворення на диплоїдному рівні. Ам. Натураліст 109: 83—92, 1976а Висновок про час виникнення деяких видів Drosophila. Природа 259: 395—396, 1976б Генетичні відмінності між новоутвореними видами. Біонаука 26: 700—701.
- Карсон, Х. Л., П. С. Нейр та Ф. М. Сене, 1975 Гібриди Drosophila в природі: Доказ обміну генами між симпатричними видами. Наука 189: 806—807.
- Карсон, Х. Л. та Х. Д. Сталкер, 1968 Взаємозв'язки політенних хромосом у гавайських видах Drosophila. II. Підгрупа D. planitibia. Університет Техасу Публікація 6818: 355-365.
- Койн, Дж. А., 1976 Відсутність генетичної схожості між двома спорідненими видами Drosophila, виявлена різними методами. Генетика 84: 593—607.
- Креддок, Е. М., 1974 Репродуктивні взаємини між гомосеквенційними видами гавайської Drosophila. Еволюція 28: 593—606.
- Креддок, Е. М. та Х. Л. Карсон, 1975 Варіабельність хромосом у ендемічному гавайському виді Drosophila. Генетика 80: s23.
- Готліб, Л. Д., 1974 Генетичне підтвердження походження Clarkia lingulata. Еволюція 2B: 244-250.
- Харді, Д. Е., 1965 Комахи Гаваїв, Т. 12. с. 814. Університет Гаваїв Прес, Гонолулу.
- Джонсон, В. Е. та Х. Л. Карсон, 1975 Алозимна варіація в Drosophila silvestris. Генетика 80: s46.
- Джонсон, В. Е., Х. Л. Карсон, К. Й. Канешіро, В. В. М. Стайнер та М. М. Купер, 1975 Генетична варіація в гавайській Drosophila. II. Алозимна диференціація в підгрупі D. planitibia. с. 563—584. У: Ізозими IV. Генетика та еволюція. Редактор: К. Л. Маркерт. Академічний прес, Нью-Йорк.
- Канешіро, К. Й., 1976 Етологічна ізоляція та філогенія в підгрупі планітібія гавайської Drosophila. Еволюція 30: 740-745.
- Канешіро, К. Й. та Ф. С. Вал, 1977 Природна гібридизація між симпатричною парою гавайської Drosophila. Ам. Натураліст (в друці).
- Левонтін, Р. С. та Дж. Л. Хаббі, 1966 Молекулярний підхід до вивчення генетичної гетерозиготності в природних популяціях. II. Кількість варіації та ступінь гетерозиготності в природних популяціях Drosophila pseudoobscura. Генетика 54: 595—609.
- Ней, М., 1972 Генетична відстань між популяціями. Ам. Натураліст 106: 283—292.
- Пауел, І. Р., 1975 Варіація білків у природних популяціях тварин. Евол. Біол. 8: 79-113.
- Саймон, Р. С., Р. С. ЛЕВОНТІН та А. А. Фццоу, 1976 Генетична гетерогенність в електрофоретичних “алелях” ксантиндегідрогенази в Drosophila pseudoobscura. Генетика 84: 609—629.
- Стейнер, В. В. М. та В. Е. Джонсон, 1973 Техніки електрофорезу гавайської Drosophila. US/IBP Острівні екосистеми IRP. Технічний звіт 30: ii + 21 с.
- Тернер Б. Дж., 1974 Генетичне розходження видів рибок з Долини Смерті: Біохімічні проти морфологічних доказів. Еволюція 18: 281-294.
- Вал, Ф. С., 1976 Генетика морфологічних відмінностей між двома інтерфертильними видами Drosophila. Генетика 83: s78.